Back Clinic Oxidative Stress Chiropractic dan Tim Pengobatan Fungsional. Stres oksidatif didefinisikan sebagai gangguan keseimbangan antara produksi oksigen reaktif (radikal bebas) dan pertahanan antioksidan. Dengan kata lain, itu adalah ketidakseimbangan antara produksi radikal bebas dan kemampuan tubuh untuk melawan atau mendetoksifikasi efek berbahaya melalui netralisasi oleh antioksidan. Stres oksidatif menyebabkan banyak kondisi patofisiologis dalam tubuh. Ini termasuk penyakit neurodegeneratif, yaitu penyakit Parkinson, penyakit Alzheimer, mutasi gen, kanker, sindrom kelelahan kronis, sindrom X rapuh, gangguan jantung dan pembuluh darah, aterosklerosis, gagal jantung, serangan jantung, dan penyakit inflamasi. Oksidasi terjadi dalam beberapa keadaan:
sel menggunakan glukosa untuk menghasilkan energi
Sistem kekebalan tubuh melawan bakteri dan menciptakan peradangan
mayat mendetoksifikasi polutan, pestisida, dan asap rokok
Ada jutaan proses yang terjadi di tubuh kita pada waktu tertentu yang dapat mengakibatkan oksidasi. Berikut adalah beberapa gejala:
Kelelahan
Kehilangan memori dan atau kabut otak
Otot dan atau nyeri sendi
Keriput disertai rambut abu-abu
Penglihatan berkurang
Sakit kepala dan kepekaan terhadap kebisingan
Kerentanan terhadap infeksi
Memilih makanan organik dan menghindari racun di lingkungan Anda membuat perbedaan besar. Ini, bersama dengan mengurangi stres, dapat bermanfaat dalam mengurangi oksidasi.
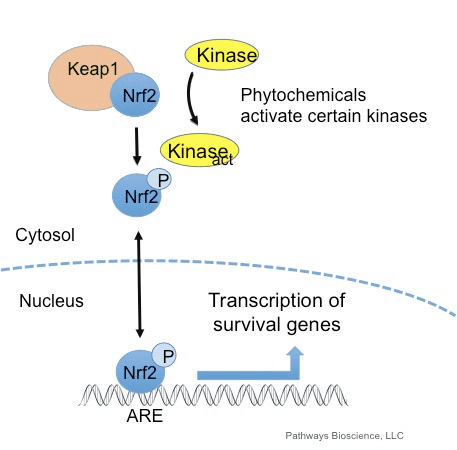
Oksidan umumnya diproduksi secara terkontrol untuk mengatur proses penting dalam tubuh manusia, termasuk pembelahan sel, peradangan, fungsi kekebalan, autophagy, dan respon stres. Namun, produksi oksidan yang tidak terkontrol ini dapat berkontribusi stres oksidatif, yang dapat mempengaruhi fungsi seluler, yang mengarah pada pengembangan toksisitas, penyakit kronis dan kanker. Mekanisme antioksidan pelindung tubuh manusia diatur oleh serangkaian jalur vital yang mengendalikan respons sel terhadap oksidan. Faktor nuklir erythroid 2 terkait faktor, atau dikenal sebagai Nrf2, adalah regulator yang muncul dari resistensi seluler terhadap oksidan. Tujuan artikel di bawah ini adalah untuk mendiskusikan dan menunjukkan peran Nrf2 yang muncul dalam fungsi mitokondria.
Abstrak
Faktor transkripsi NF-E2 p45-related factor 2 (Nrf2; nama gen NFE2L2) memungkinkan adaptasi dan kelangsungan hidup dalam kondisi stres dengan mengatur ekspresi gen dari beragam jaringan protein sitoprotektif, termasuk enzim antioksidan, anti-inflamasi, dan detoksifikasi sebagai protein yang membantu dalam perbaikan atau pembuangan makromolekul yang rusak. Nrf2 memiliki peran penting dalam pemeliharaan homeostasis redoks seluler dengan mengatur biosintesis, pemanfaatan, dan regenerasi glutathione, thioredoxin, dan NADPH dan dengan mengontrol produksi spesies oksigen reaktif oleh mitokondria dan NADPH oksidase. Dalam kondisi homeostatis, Nrf2 mempengaruhi potensi membran mitokondria, oksidasi asam lemak, ketersediaan substrat (NADH dan FADH2 / suksinat) untuk respirasi, dan sintesis ATP. Dalam kondisi stres atau stimulasi faktor pertumbuhan, aktivasi Nrf2 melawan peningkatan produksi spesies oksigen reaktif di mitokondria melalui peningkatan transkripsi dari protein 3 yang tidak digabungkan dan mempengaruhi biogenesis mitokondria dengan mempertahankan tingkat faktor pernapasan inti 1 dan reseptor yang diaktifkan proliferator peroksisom? koaktivator 1 ?, serta dengan mempromosikan biosintesis nukleotida purin. Aktivator farmakologis Nrf2, seperti sulforaphane isothiocyanate yang terjadi secara alami, menghambat pembukaan yang dimediasi oksidan pada pori transisi permeabilitas mitokondria dan pembengkakan mitokondria. Anehnya, senyawa sintetik 1,4-difenil-1,2,3-triazol, awalnya dirancang sebagai aktivator Nrf2, ditemukan mempromosikan mitofagi, sehingga berkontribusi pada homeostasis mitokondria secara keseluruhan. Jadi, Nrf2 adalah pemain penting dalam mendukung integritas struktural dan fungsional mitokondria, dan peran ini sangat penting dalam kondisi stres.
Kata kunci:Bioenergetika, Sitoproteksi, Keap1, Mitochondria, Nrf2, Radikal bebas
Highlight
Nrf2 memiliki peran penting dalam menjaga homeostasis sel redoks.
Nrf2 mempengaruhi potensi membran mitokondria dan sintesis ATP.
Nrf2 mempengaruhi oksidasi asam lemak mitokondria.
Nrf2 mendukung integritas struktural dan fungsional mitokondria.
Aktivator Nrf2 memiliki efek menguntungkan ketika fungsi mitokondria dikompromikan.
Pengantar
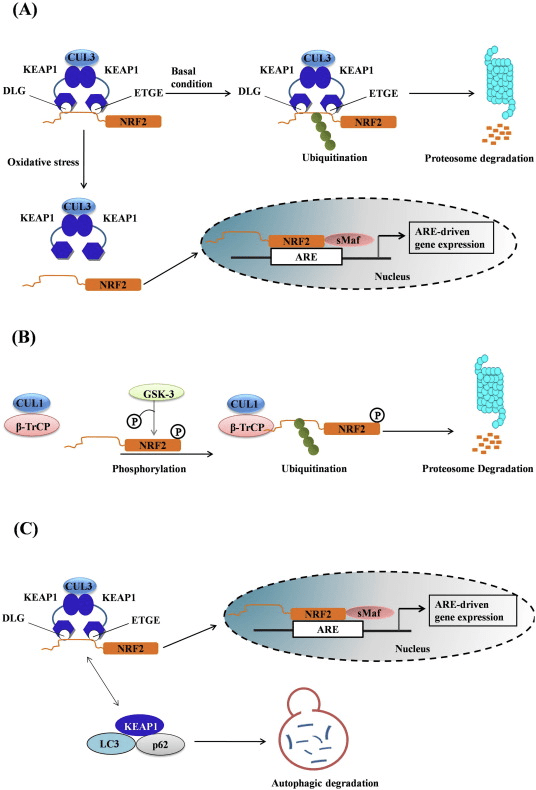
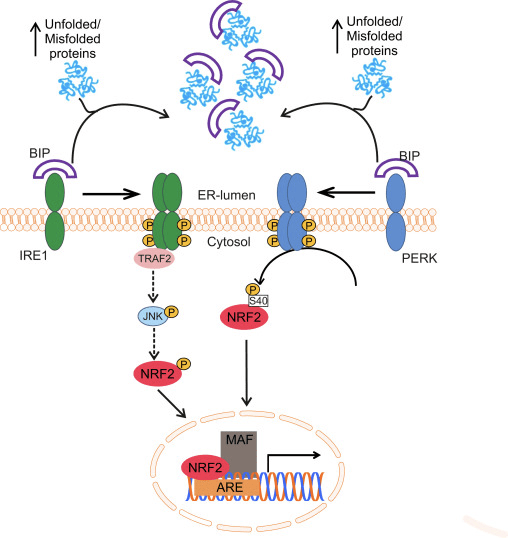
Faktor transkripsi NF-E2 p45 terkait faktor 2 (Nrf2; nama gen NFE2L2) mengatur ekspresi jaringan gen yang mengkode protein dengan beragam aktivitas sitoprotektif. Nrf2 sendiri dikendalikan terutama pada tingkat kestabilan protein. Di bawah kondisi basal, Nrf2 adalah protein berumur pendek yang mengalami ubiquitination terus menerus dan degradasi proteasomal. Ada tiga sistem ligase ubiquitin yang berkontribusi terhadap degradasi Nrf2. Secara historis, regulator negatif pertama Nrf2 yang ditemukan adalah protein terkait ECH seperti Kelch 1 (Keap1) [1], protein adaptor substrat untuk Cullin 3 (Cul3) / Rbx1 ubiquitin ligase [2], [3], [ 4]. Keap1 menggunakan mekanisme siklik yang sangat efisien untuk menargetkan Nrf2 untuk ubiquitination dan degradasi proteasomal, di mana Keap1 terus diregenerasi, memungkinkan siklus untuk dilanjutkan (Gbr. 1A) [5]. Nrf2 juga mengalami degradasi yang dimediasi oleh glikogen sintase kinase (GSK) 3 /? - Ligase ubiquitin berbasis Cul1 yang bergantung pada TrCP [6], [7]. Baru-baru ini, dilaporkan bahwa, selama kondisi stres retikulum endoplasma, Nrf2 ada di mana-mana dan terdegradasi dalam proses yang dimediasi oleh E3 ubiquitin ligase Hrd1 [8].
Gambar 1 Pengikatan sekuensial siklik dan model regenerasi untuk degradasi Nrf1 yang dimediasi oleh Keap2. (A) Nrf2 mengikat secara berurutan ke dimer Keap1 gratis: pertama melalui domain berikatan tinggi ETGE (tongkat merah) dan kemudian melalui domain pengikatan DLG (tongkat hitam) afinitas rendah. Dalam konformasi kompleks protein ini, Nrf2 mengalami ubiquitination dan ditargetkan untuk degradasi proteasomal. Keap1 gratis diregenerasi dan dapat berikatan dengan Nrf2 yang baru diterjemahkan, dan siklusnya dimulai lagi. (B) Inducer (berlian putih) bereaksi dengan cysteines sensor Keap1 (tongkat biru), yang mengarah ke perubahan konformasi dan gangguan aktivitas adaptor substrat. Keap1 gratis tidak diregenerasi, dan Nrf2 yang baru disintesis berakumulasi dan mentranslokasi ke nukleus.
Selain berfungsi sebagai protein adapter substrat ubiquitin ligase, Keap1 juga merupakan sensor untuk beragam aktivator molekul kecil Nrf2 (disebut inducers) [9]. Inducer memblokir siklus degradasi Nrf1 yang dimediasi oleh Keap2 dengan memodifikasi residu sistein spesifik secara kimia dalam Keap1 [10], [11] atau dengan langsung mengganggu antarmuka antarmuka Keap1: Nrf2 [12], [13]. Akibatnya, Nrf2 tidak terdegradasi, dan faktor transkripsi mengakumulasi dan mentranslokasi ke nukleus (Gambar 1B), di mana ia membentuk heterodimer dengan protein Maf kecil; berikatan dengan unsur-unsur respon antioksidan, daerah pengatur hulu dari gen targetnya; dan memulai transkripsi [14], [15], [16]. Baterai target Nrf2 terdiri dari protein dengan beragam fungsi sitoprotektif, termasuk enzim metabolisme xenobiotik, protein dengan fungsi antioksidan dan anti-inflamasi, dan subunit proteasomal, serta protein yang mengatur homeostasis sel redoks dan berpartisipasi dalam metabolisme perantara.
Nrf2: Master Regulator Homeostasis Redoks Seluler
Fungsi Nrf2 sebagai pengatur utama homeostasis redoks seluler sudah dikenal luas. Ekspresi gen dari kedua katalitik dan subunit pengatur dari? -Glutamyl cysteine ligase, enzim yang mengkatalis langkah pembatas laju dalam biosintesis glutathione tereduksi (GSH), secara langsung diatur oleh Nrf2 [17]. Subunit xCT dari sistem xc-, yang mengimpor sistin ke dalam sel, juga merupakan target transkripsi langsung dari Nrf2 [18]. Di dalam sel, sistin mengalami konversi menjadi sistein, prekursor untuk biosintesis GSH. Selain perannya dalam biosintesis GSH, Nrf2 menyediakan sarana untuk pemeliharaan glutathione dalam keadaan tereduksi dengan regulasi transkripsi terkoordinasi dari glutathione reduktase 1 [19], [20], yang mengurangi glutathione teroksidasi menjadi GSH menggunakan pengurangan ekuivalen dari NADPH . NADPH yang dibutuhkan disediakan oleh empat enzim penghasil NADPH utama, enzim malat 1 (ME1), isocitrate dehydrogenase 1 (IDH1), glukosa-6-fosfat dehidrogenase (G6PD), dan 6-fosfoglukonat dehidrogenase (PGD), yang semuanya adalah secara transkripsi diatur sebagian oleh Nrf2 (Gbr. 2) [21], [22], [23], [24]. Anehnya, Nrf2 juga mengatur ekspresi gen yang dapat diinduksi dari bentuk sitosol, mikrosomal, dan mitokondria dari aldehida dehidrogenase [25], yang menggunakan NAD (P) + sebagai kofaktor, sehingga menimbulkan NAD (P) H. Memang, tingkat NADPH dan rasio NADPH / NADP + lebih rendah pada fibroblas embrionik yang diisolasi dari tikus Nrf2-knockout (Nrf2-KO) dibandingkan dengan sel dari rekan tipe liar (WT) mereka, dan tingkat NADPH menurun saat Nrf2 knockdown di garis sel kanker dengan Nrf2 aktif secara konstitutif [26]. Seperti yang diharapkan, tingkat GSH lebih rendah dalam sel di mana Nrf2 telah terganggu; sebaliknya, aktivasi Nrf2 dengan cara genetik atau farmakologis menyebabkan peningkatan regulasi GSH [27]. Yang penting, Nrf28 juga mengatur ekspresi gen tioredoksin [29], [2], [30], tioredoksin reduktase 31 [32], [1], [28], [29], dan sulfiredoksin [32], yang penting untuk reduksi tiol protein teroksidasi.
Gambar 2 Peran Nrf2 dalam metabolisme sel yang berproliferasi cepat. Nrf2 adalah regulator positif dari gen yang mengkode enzim di kedua lengan oksidatif [yaitu, glukosa-6-fosfat dehidrogenase (G6PD) dan 6-fosfoglukonat dehidrogenase (PGD)] dan lengan nonoksidatif [yaitu, transaldolase 1 (TALDO1) dan transketolase ( TKT)] dari jalur pentosa fosfat. G6PD dan PGD menghasilkan NADPH. Nrf2 juga mengatur ekspresi gen dari dua enzim penghasil NADPH lainnya, enzim malat 1 (ME1) dan isocitrate dehydrogenase 1 (IDH1). Ekspresi gen fosforibosil pirofosfat amidotransferase (PPAT), yang mengkatalisis masuk ke jalur biosintetik purin de novo, juga diatur secara positif oleh Nrf2, seperti ekspresi methylenetetrahydrofolate dehydrogenase 2 (MTHFD2), enzim mitokondria dengan peran penting dalam menyediakan unit satu karbon untuk biosintesis purin de novo. Piruvat kinase (PK) diatur secara negatif oleh Nrf2 dan diharapkan mendukung penumpukan zat antara glikolitik dan, bersama dengan G6PD, saluran metabolit melalui jalur pentosa fosfat dan sintesis asam nukleat, asam amino, dan fosfolipid. Nrf2 secara negatif mengatur ekspresi gen ATP-sitrat lyase (CL), yang dapat meningkatkan ketersediaan sitrat untuk penggunaan mitokondria atau (melalui isocitrate) untuk IDH1. Merah dan biru masing-masing menunjukkan regulasi positif dan negatif. Mitokondria ditampilkan dalam warna abu-abu. Singkatan metabolit: G-6-P, glukosa 6-fosfat; F-6-P, fruktosa 6-fosfat; F-1,6-BP, fruktosa 1,6-bifosfat; GA-3-P, gliseraldehida 3-fosfat; 3-PG, 3-fosfogliserat; PEP, fosfoenolpiruvat; 6-P-Gl, 6-fosfoglukonolakton; 6-PG, 6-fosfoglukonat; R-5-P, ribulosa 5-fosfat; PRPP, 5-fosforibosil -? - 1-pirofosfat; THF, tetrahidrofolat; IMP, inosin monofosfat; AMP, adenosin monofosfat; GMP, guanosine monophosphate.
Mengingat peran penting Nrf2 sebagai pengatur utama homeostasis sel redoks, tidak mengherankan bahwa, dibandingkan dengan sel WT, tingkat spesies oksigen reaktif (ROS) lebih tinggi dalam sel di mana Nrf2 telah terganggu (Nrf2-KO) [35]. Perbedaan ini sangat mencolok pada tantangan dengan agen yang menyebabkan stres oksidatif. Selain itu, sel kekurangan Nrf2 jauh lebih sensitif terhadap toksisitas oksidan dari berbagai jenis dan tidak dapat dilindungi oleh induser Nrf2, yang, dalam kondisi yang sama, memberikan perlindungan yang efisien dan tahan lama untuk sel WT [29], [36] , [37]. Selain homeostasis redoks selular secara keseluruhan, Nrf2 juga penting untuk pemeliharaan homeostasis mitokondria redoks. Jadi, dibandingkan dengan WT, total NADH mitokondria meningkat secara signifikan di Keap1-KO dan secara dramatis menurun pada sel Nrf2-KO [35].
Dengan menggunakan pencitraan sel hidup, kami baru-baru ini memantau tingkat produksi ROS di kokultur glioneuronal primer dan irisan jaringan otak yang diisolasi dari tikus WT, Nrf2-KO, atau Keap1-knockdown (Keap1-KD) [38]. Seperti yang diharapkan, tingkat produksi ROS lebih cepat di sel Nrf2-KO dan jaringan dibandingkan dengan rekan-rekan WT mereka. Namun, kami membuat pengamatan tak terduga bahwa, dibandingkan dengan WT, sel Keap1-KD juga memiliki tingkat produksi ROS yang lebih tinggi, meskipun besarnya perbedaan antara WT dan genotipe Keap1-KD lebih kecil daripada antara WT dan Nrf2-KO . Kami kemudian menganalisis tingkat mRNA NOX2 dan NOX4, subunit katalitik dari dua isoform NADPH oksidase (NOX) yang telah terlibat dalam patologi otak, dan menemukan bahwa NOX2 secara dramatis meningkat pada kondisi defisiensi Nrf2, sedangkan NOX4 diregulasi saat Nrf2 diaktifkan secara konstitutif, meskipun pada tingkat yang lebih kecil. Secara kuantitatif, besarnya peningkatan regulasi dalam sel dan jaringan dari tikus mutan sejajar dengan peningkatan yang sesuai dalam produksi ROS [38]. Menariknya, tidak hanya Nrf2 mengatur NADPH oksidase, tetapi ROS yang diproduksi oleh NADPH oksidase dapat mengaktifkan Nrf2, seperti yang ditunjukkan pada sel-sel epitel paru dan kardiomiosit [39], [40]. Selain itu, studi yang sangat baru-baru ini telah menunjukkan bahwa NADPH oksidase-tergantung aktivasi Nrf2 merupakan mekanisme endogen penting untuk perlindungan terhadap kerusakan mitokondria dan kematian sel di jantung selama overload tekanan kronis [41].
Selain aktivitas katalitik NADPH oksidase, respirasi mitokondria merupakan sumber intraseluler utama lainnya dari ROS. Dengan menggunakan mitokondria-spesifik probe MitoSOX, kami telah memeriksa kontribusi ROS asal mitokondria terhadap produksi ROS secara keseluruhan pada kokultur glioneuronal primer yang diisolasi. dari WT, Nrf2-KO, atau tikus Keap1-KD [38]. Seperti yang diharapkan, sel Nrf2-KO memiliki tingkat produksi mitosondria ROS lebih tinggi daripada WT. Sesuai dengan temuan untuk produksi ROS secara keseluruhan, tingkat produksi ROS mitokondria di Keap1-KD juga lebih tinggi dibandingkan dengan sel WT. Yang penting, pemblokiran kompleks I dengan rotenone menyebabkan peningkatan dramatis dalam produksi ROS mitokondria pada sel WT dan Keap1-KD, tetapi tidak memiliki efek pada sel Nrf2-KO. Berbeda dengan peningkatan yang diharapkan dalam produksi ROS mitokondria pada sel WT setelah penambahan piruvat (untuk meningkatkan ketersediaan NADH, meningkatkan potensi membran mitokondria, dan menormalkan respirasi), produksi ROS menurun pada sel Nrf2-KO. Bersama-sama, temuan ini sangat menunjukkan bahwa, dengan tidak adanya Nrf2: (i) aktivitas kompleks I terganggu, (ii) gangguan aktivitas kompleks I adalah karena keterbatasan substrat, dan (iii) gangguan aktivitas kompleks. Saya adalah salah satu alasan utama untuk peningkatan produksi ROS mitokondria, mungkin karena arus balik elektron dari kompleks II.
Nrf2 Mempengaruhi Potensi dan Respirasi Membran Mitokondria
Potensi membran mitokondria (?? m) adalah indikator universal kesehatan mitokondria dan keadaan metabolisme sel. Dalam sel yang sehat, m dipelihara oleh rantai pernapasan mitokondria. Menariknya, pelabelan isotop yang stabil dengan asam amino dalam studi proteomik berbasis kultur dalam garis sel MCF10A epitel payudara manusia nontumorigenik reseptor estrogen-negatif telah menunjukkan bahwa komponen rantai transpor elektron mitokondria NDUFA4 diregulasi oleh aktivasi farmakologis (oleh sulforaphane) dari Nrf2, sedangkan peningkatan regulasi genetik Nrf2 (oleh knockdown Keap1) menyebabkan regulasi turun subunit oksidase sitokrom c COX2 dan COX4I1 [42]. Sebuah studi tentang proteome hati menggunakan elektroforesis gel dua dimensi dan spektrometri massa desorpsi / ionisasi laser yang dibantu matriks telah menemukan bahwa Nrf2 mengatur ekspresi subunit sintase ATP? [43]. Selain itu, protein mitokondria DJ-1, yang berperan dalam pemeliharaan aktivitas kompleks I [44], telah dilaporkan untuk menstabilkan Nrf2 [45], [46], meskipun efek neuroprotektif dari aktivasi farmakologis atau genetik dari Nrf2 tidak bergantung pada DJ-1 [47]. Namun, konsekuensi dari pengamatan terhadap fungsi mitokondria ini belum diselidiki.
Sesuai dengan aktivitas gangguan kompleks I dalam kondisi defisiensi Nrf2, basal m lebih rendah pada fibroblas embrionik tikus Nrf2-KO (MEF) dan sel glioneuronal primer yang dikultur dibandingkan dengan rekan WT mereka (Gbr. 3, sisipan) [35]. Sebaliknya, basal m lebih tinggi ketika Nrf2 secara genetik diatur secara konstitutif (dengan knockdown atau knockout dari Keap1). Perbedaan m di antara genotipe menunjukkan bahwa respirasi dipengaruhi oleh aktivitas Nrf2. Memang, evaluasi konsumsi oksigen dalam keadaan basal telah mengungkapkan bahwa, dibandingkan dengan WT, konsumsi oksigen lebih rendah pada MEF Nrf2-KO dan Keap1-KO, masing-masing sebesar ~ 50 dan ~ 35%.
Gambar 3 Mengusulkan mekanisme untuk fungsi mitokondria yang dikompromikan dalam kondisi defisiensi Nrf2. (1) Penurunan kadar ME1, IDH1, G6PD, dan PGD mengakibatkan penurunan kadar NADPH. (2) Tingkat GSH juga rendah. (3) Aktivitas ME1 yang rendah dapat menurunkan kumpulan piruvat yang memasuki mitokondria. (4) Pembangkitan NADH lebih lambat, menyebabkan gangguan aktivitas kompleks I dan peningkatan produksi ROS mitokondria. (5) Reduksi FAD menjadi FADH2 dalam protein mitokondria juga menurun, menurunkan aliran elektron dari FADH2 ke UbQ dan menjadi kompleks III. (6) Pembentukan UbQH2 yang lebih lambat dapat menurunkan aktivitas enzim dari suksinat dehidrogenase. (7) Peningkatan kadar ROS selanjutnya dapat menghambat aktivitas kompleks II. (8) Efisiensi oksidasi asam lemak yang lebih rendah berkontribusi pada penurunan ketersediaan substrat untuk respirasi mitokondria. (9) Glikolisis ditingkatkan sebagai mekanisme kompensasi untuk penurunan produksi ATP dalam fosforilasi oksidatif. (10) ATP sintase beroperasi secara terbalik untuk mempertahankan m. Merah dan biru masing-masing menunjukkan regulasi naik dan turun. Kotak tersebut menandakan ketersediaan bukti eksperimental. Inset tersebut menunjukkan gambar mitokondria dari astrosit kortikal WT dan Nrf2-KO yang divisualisasikan oleh probe fluoresen potensiometri tetramethylrhodamine methyl ester (TMRM; 25 nM). Bilah skala, 20 m.
Perbedaan dalam m dan respirasi antar genotipe ini dicerminkan oleh laju pemanfaatan substrat untuk respirasi mitokondria. Penerapan substrat untuk siklus asam trikarboksilat (TCA) (malat / piruvat, yang pada gilirannya meningkatkan produksi substrat kompleks I NADH) atau metil suksinat, substrat untuk kompleks II, menyebabkan peningkatan bertahap dalam m di kedua WT dan neuron Keap1-KD, tetapi laju peningkatannya lebih tinggi pada sel Keap1-KD. Lebih penting lagi, bentuk respons terhadap siklus substrat TCA ini berbeda antara dua genotipe, di mana peningkatan cepat m dalam sel Keap1-KD setelah penambahan substrat diikuti dengan penurunan cepat daripada dataran tinggi, menunjukkan penurunan yang tidak biasa. konsumsi media yang cepat. Temuan ini sejalan dengan kadar malat, piruvat, dan suksinat yang jauh lebih rendah (sebesar 50-70%) yang telah diamati setelah denyut nadi [U-1C13] glukosa 6 jam di Keap1-KO dibandingkan dengan WT MEF. sel [24]. Pada neuron Nrf2-KO, hanya piruvat yang mampu meningkatkan m, sedangkan malat dan metil suksinat menyebabkan depolarisasi ringan. Pengaruh Nrf2 pada produksi substrat mitokondria tampaknya menjadi mekanisme utama dimana Nrf2 mempengaruhi fungsi mitokondria. Indeks redoks NADH mitokondria (keseimbangan antara konsumsi NADH oleh kompleks I dan produksi NADPH dalam siklus TCA) secara signifikan lebih rendah dalam sel Nrf2-KO dibandingkan dengan rekan WT mereka, dan lebih jauh lagi, tingkat regenerasi kumpulan dari NADH dan FADH2 setelah penghambatan kompleks IV (dengan menggunakan NaCN) lebih lambat pada sel mutan.
Dalam mitokondria yang diisolasi dari otak dan hati murine, suplementasi substrat untuk kompleks I atau kompleks II meningkatkan laju konsumsi oksigen lebih kuat ketika Nrf2 diaktifkan dan kurang efisien ketika Nrf2 terganggu [35]. Jadi, malat menginduksi tingkat konsumsi oksigen yang lebih tinggi pada Keap1-KD dibandingkan dengan WT, tetapi efeknya lebih lemah pada mitokondria Nrf2-KO. Demikian pula, dengan adanya rotenon (ketika kompleks I dihambat), suksinat mengaktifkan konsumsi oksigen ke tingkat yang lebih besar di Keap1-KD dibandingkan dengan WT, sedangkan respons dalam mitokondria Nrf2-KO berkurang. Selain itu, kultur neuron primer Nrf2-KO dan tikus lebih sensitif terhadap toksisitas kompleks II inhibitor asam 3-nitropropionat dan malonat kompleks, sedangkan transplantasi intrastriatal dari astrosit yang mengekspresikan Nrf2-overexpressing bersifat protektif [48]. Demikian pula, tikus Nrf49-KO lebih sensitif terhadap, sedangkan aktivasi genetik atau farmakologis Nrf2 memiliki efek perlindungan terhadap, neurotoksisitas yang disebabkan oleh penghambat I kompleks 2-metil-1-fenilpiridinium ion dalam 4-metil-1-fenil-4, Model hewan 1,2,3,6-tetrahidropiridin penyakit Parkinson [49], [50], [51], [52], [53], [54], [55], [56], [57], [58], [59], [60], [61].
Rasio kontrol pernapasan (RCR), rasio Negara 3 (terstimulasi ADP) dengan respirasi keadaan 4 (tidak ada ADP), menurun dengan tidak adanya Nrf2, tetapi RCR serupa antara Keap1-KD dan mitokondria WT [35 ]. Karena RCR merupakan indikasi derajat kopling aktivitas rantai pernapasan mitokondria dengan fosforilasi oksidatif, temuan ini menunjukkan bahwa laju respirasi yang lebih tinggi dalam mitokondria Keap1-KD bukan karena pelepasan fosforilasi oksidatif. Lebih lanjut menunjukkan bahwa fosforilasi oksidatif lebih efisien ketika Nrf2 diaktifkan. Laju respirasi yang lebih tinggi di mitokondria Keap1-KD konsisten dengan tingkat produksi ROS mitokondria yang lebih tinggi [38] karena laju respirasi yang lebih tinggi dapat menyebabkan peningkatan kebocoran elektron. Namun, dalam kondisi stres oksidatif, peningkatan produksi ROS diimbangi oleh regulasi transkripsi yang bergantung pada Nrf2 dari uncoupling protein 3 (UCP3), yang meningkatkan konduktansi proton dari membran dalam mitokondria dan akibatnya menurunkan produksi superoksida [62]. Baru-baru ini, ditunjukkan bahwa produk peroksidasi lipid 4-hidroksi-2-nonenal memediasi peningkatan regulasi UCP2 yang bergantung pada Nrf3 dalam kardiomiosit; ini mungkin sangat penting untuk perlindungan di bawah kondisi stres oksidatif seperti selama reparfusi iskemia [63].
Nrf2 Mempengaruhi Efisiensi Fosforilasi Oksidatif dan Sintesis ATP
Sesuai dengan efek Nrf2 pada respirasi, dalam mitokondria otak dan hati, defisiensi Nrf2 mengakibatkan penurunan efisiensi fosforilasi oksidatif (seperti yang diperkirakan dengan rasio ADP terhadap oksigen, yang dikonsumsi untuk sintesis ATP), sedangkan aktivasi Nrf2 (Keap1) -KD) memiliki efek sebaliknya [35]. Dibandingkan dengan WT, tingkat ATP secara signifikan lebih tinggi dalam sel dengan peningkatan regulasi konstitutif Nrf2 dan lebih rendah ketika Nrf2 dirobohkan [64] atau terganggu [35]. Lebih lanjut, penggunaan inhibitor fosforilasi oksidatif (oligomisin) atau glikolisis (asam iodoasetat) telah mengungkapkan bahwa Nrf2 mengubah cara sel memproduksi ATP. Jadi, dalam neuron WT, oligomisin menyebabkan penurunan total ATP dan asam iodoasetat tidak berpengaruh lebih lanjut. Hebatnya, dalam sel Nrf2-KO, oligomisin meningkatkan kadar ATP, yang kemudian perlahan, tetapi seluruhnya, habis oleh asam iodoasetat, menunjukkan bahwa dengan tidak adanya Nrf2, glikolisis, dan bukan fosforilasi oksidatif, adalah sumber utama produksi ATP. Menariknya, meskipun terjadi peningkatan efisiensi fosforilasi oksidatif dalam sel Keap1-KD, penambahan oligomisin menghasilkan penurunan ~ 80% tingkat ATP, dan asam iodoasetat menyebabkan penurunan ~ 20% lebih lanjut. Jadi, baik defisiensi Nrf2 atau aktivasi konstitutifnya mengurangi kontribusi fosforilasi oksidatif dan meningkatkan kontribusi glikolisis terhadap sintesis ATP. Efek ini terutama diucapkan ketika Nrf2 tidak ada dan konsisten dengan ketergantungan m pada keberadaan glukosa dalam medium [35] dan peningkatan kadar zat antara glikolitik (G-6-P, F-6-P , dihydroxyacetone fosfat, piruvat, dan laktat) setelah knockdown Nrf2 [24].
Peningkatan kadar ATP setelah penghambatan F1F0-ATPase oleh oligomisin menunjukkan bahwa dengan tidak adanya Nrf2, F1F0-ATPase berfungsi sebagai ATPase dan bukan sintase ATP, yaitu beroperasi secara terbalik. Pembalikan aktivitas seperti itu kemungkinan besar mencerminkan kebutuhan untuk memompa proton melintasi membran mitokondria bagian dalam dalam upaya untuk mempertahankan m, yang sangat penting untuk integritas fungsional organel ini. Pembalikan fungsi F1F0-ATPase juga dibuktikan oleh depolarisasi mitokondria yang diamati pada pemberian oligomisin ke sel Nrf2-KO, yang sangat kontras dengan hiperpolarisasi yang terjadi pada rekan-rekan mereka yang kekurangan WT atau Keap1 [35]. Secara keseluruhan, tampaknya dalam kondisi defisiensi Nrf2, ATP diproduksi terutama dalam glikolisis, dan ATP ini kemudian digunakan sebagian oleh F1F0-ATPase untuk mempertahankan m.
Nrf2 Meningkatkan Oksidasi Asam Lemak Mitokondria
Efek defisiensi Nrf2 pada m ?? terutama terlihat ketika sel diinkubasi dalam medium tanpa glukosa, dan m ~ 50% lebih rendah pada Nrf2-KO dibandingkan dengan sel WT [35]. Dalam kondisi kekurangan glukosa, oksidasi asam lemak mitokondria (FAO) adalah penyedia utama substrat untuk respirasi dan fosforilasi oksidatif, menunjukkan bahwa Nrf2 dapat mempengaruhi FAO. Memang, efisiensi FAO untuk asam palmitat asam lemak jenuh rantai panjang (C16: 0) dan asam heksanoat rantai pendek (C6: 0) lebih tinggi pada MEF Keap1-KO dan mitokondria jantung dan hati yang terisolasi daripada di Rekan WT, sedangkan itu lebih rendah pada sel Nrf2-KO dan mitokondria [65]. Efek ini juga sangat relevan bagi manusia: memang, perubahan metabolik yang mengindikasikan integrasi FAO yang lebih baik dengan aktivitas siklus TCA telah dilaporkan terjadi pada studi intervensi manusia dengan diet kaya glukorafanin, pendahulu sulforaphane aktivator Nrf2 klasik [ 66].
Selama langkah pertama FAO mitokondria, hidrogen pro-R dari daun karbona sebagai hidrida yang mengurangi kofaktor FAD menjadi FADH2, yang pada gilirannya mentransfer elektron ke ubikuinon (UbQ) dalam rantai pernapasan, yang pada akhirnya berkontribusi pada produksi ATP. . Sedangkan stimulasi FAO oleh palmitoylcarnitine dengan tidak adanya glukosa menyebabkan peningkatan yang diharapkan dalam kadar ATP dalam sel WT dan Keap1-KO, dengan peningkatan ATP yang lebih cepat dalam sel Keap1-KO, pengobatan yang sama tidak menghasilkan perubahan ATP pada Nrf2-KO. MEF [65]. Percobaan ini menunjukkan bahwa, dengan tidak adanya Nrf2, FAO ditekan, dan lebih jauh, itu berimplikasi penekanan FAO sebagai salah satu alasan untuk tingkat ATP yang lebih rendah dalam kondisi defisiensi Nrf2 [35].
Khususnya, sel 293 T manusia di mana Nrf2 telah dibungkam memiliki ekspresi yang lebih rendah dari CPT1 dan CPT2 [67], dua isoform karnitin palmitoyltransferase (CPT), enzim pembatas laju di mitochondrial FAO. Dalam kesepakatan, tingkat mRNA Cpt1 lebih rendah di hati Nrf2-KO dibandingkan dengan tikus WT [68]. CPT mengkatalisis transfer gugus asil dari asil lemak rantai panjang dari koenzim A ke l-karnitin dan dengan demikian memungkinkan pengimporan acylcarnitine dari sitoplasma ke mitokondria. Meskipun ini belum diperiksa hingga saat ini, ada kemungkinan bahwa selain efek transkripsi pada ekspresi CPT1, Nrf2 juga dapat mempengaruhi fungsi enzim ini dengan mengontrol tingkat inhibitor alosterik utamanya, malonyl-CoA. Hal ini karena, oleh mekanisme yang saat ini tidak jelas, Nrf2 mengatur secara negatif ekspresi stearoyl CoA desaturase (SCD) [69] dan lyase sitrat (CL) [69], [70]. Anehnya, KO atau penghambatan SCD menyebabkan peningkatan fosforilasi dan aktivasi AMP-activated protein kinase (AMPK) [71], [72], [73], dan dapat dispekulasikan bahwa, dengan tidak adanya Nrf2, tingkat SCD akan meningkat, yang pada gilirannya menurunkan aktivitas AMPK. Hal ini dapat semakin diperparah oleh berkurangnya kadar protein AMPK yang telah diamati pada hati tikus Nrf2-KO [68], sebuah temuan yang sesuai dengan peningkatan kadar AMPK, yang telah dilaporkan dalam hati Keap1-KD tikus [74]. Salah satu konsekuensi dari penurunan aktivitas AMPK adalah menghilangkan fosforilasi penghambatannya (pada Ser79) dari asetil-KoA karboksilase (ACC) [75], yang dapat ditranskripsikan secara lebih lanjut dengan tidak adanya Nrf2 karena ini diturunkan oleh aktivasi Nrf2 [70 ]. Aktivitas ACC yang tinggi, dalam kombinasi dengan ekspresi CL yang diregulasi yang akan meningkatkan produksi asetil-CoA, substrat untuk ACC, akhirnya dapat meningkatkan tingkat produk ACC, malonyl-CoA. Tingginya kadar malonyl-CoA akan menghambat CPT, sehingga mengurangi pengangkutan asam lemak ke mitokondria. Akhirnya, Nrf2 secara positif mengatur ekspresi CD36 [76], sebuah translocase yang mengimpor asam lemak melintasi plasma dan membran mitokondria. Dengan demikian, satu mekanisme dimana Nrf2 dapat mempengaruhi efisiensi FAO mitokondria adalah dengan mengatur impor asam lemak rantai panjang ke mitokondria.
Selain regulasi transkripsi langsung, Nrf2 juga dapat mengubah efisiensi FAO mitokondria oleh efeknya pada metabolisme redoks seluler. Ini mungkin sangat relevan ketika aktivitas Nrf2 rendah atau tidak ada, kondisi yang menggeser status redoks seluler menuju keadaan teroksidasi. Memang, beberapa enzim FAO telah diidentifikasi sebagai sensitif terhadap perubahan redoks. Salah satu enzim tersebut adalah rantai panjang asil-CoA dehidrogenase (VLCAD), yang menyumbang lebih dari 80% ke aktivitas dehidrogenasi palmitoyl-CoA dalam jaringan manusia [77]. Menariknya, Hurd et al. [78] telah menunjukkan bahwa VLCAD mengandung residu sistein yang secara signifikan mengubah keadaan redoks mereka setelah terpapar mitokondria jantung tikus terisolasi ke H2O2. Selain itu, S-nitrosilasi VCAD hepatik murin di Cys238 meningkatkan efisiensi katalitik enzim [79], dan kemungkinan oksidasi sistein yang sama mungkin memiliki efek sebaliknya, akhirnya menurunkan efisiensi mitokondria FAO. Oleh karena itu mungkin bahwa, meskipun tingkat ekspresi VLCAD tidak berbeda secara signifikan dalam WT, Nrf2-KO, atau Keap1-KO MEF [65], aktivitas enzim VLCAD bisa lebih rendah tanpa adanya Nrf2 karena tingkat yang lebih tinggi. dari ROS.
Berdasarkan semua temuan ini, dapat diusulkan bahwa (Gbr. 3): dengan tidak adanya Nrf2, tingkat NADPH lebih rendah karena penurunan ekspresi ME1, IDH1, G6PD, dan PGD. Kadar glutathione yang tereduksi juga lebih rendah karena penurunan ekspresi enzim yang berpartisipasi dalam biosintesis dan regenerasi dan tingkat NADPH yang lebih rendah yang diperlukan untuk konversi yang teroksidasi menjadi bentuk glutathione yang tereduksi. Ekspresi ME1 yang rendah akan mengurangi kumpulan piruvat yang memasuki mitokondria, dengan glikolisis menjadi sumber utama piruvat. Pembangkitan NADH lebih lambat, menyebabkan gangguan aktivitas kompleks I dan peningkatan produksi ROS mitokondria. Reduksi FAD menjadi FADH2 juga lebih lambat, setidaknya sebagian karena oksidasi asam lemak yang kurang efisien, mengganggu aliran elektron dari FADH2 ke UbQ dan ke kompleks III. Karena UbQH2 adalah aktivator dari suksinat dehidrogenase [80], memperlambat pembentukannya dapat menurunkan aktivitas enzim dari suksinat dehidrogenase. Peningkatan kadar superoksida dan hidrogen peroksida dapat menghambat aktivitas kompleks II lebih lanjut [81]. Efisiensi oksidasi asam lemak yang lebih rendah berkontribusi pada penurunan ketersediaan substrat untuk respirasi mitokondria dan produksi ATP dalam fosforilasi oksidatif. Sebagai mekanisme kompensasi, glikolisis ditingkatkan. ATP sintase berfungsi secara terbalik, sebagai ATPase, dalam upaya untuk mempertahankan ?? m.
Nrf2 dan Mitokondria Biogenesis
Telah dilaporkan bahwa, dibandingkan dengan WT, hati mencit Nrf2-KO memiliki kandungan mitokondria yang lebih rendah (sebagaimana ditentukan oleh rasio mitokondria terhadap DNA inti); ini selanjutnya menurun dengan cepat 24 jam pada tikus WT dan Nrf2-KO; sebaliknya, meskipun tidak berbeda dengan WT dalam kondisi makan normal, kandungan mitokondria pada tikus dengan aktivitas Nrf2 tinggi tidak dipengaruhi oleh puasa [82]. Menariknya, suplementasi dengan aktivator Nrf2 (R) -? - asam lipoat [83], [84], [85] meningkatkan biogenesis mitokondria dalam adiposit 3T3-L1 [86]. Dua kelas regulator transkripsi nuklir memainkan peran penting dalam biogenesis mitokondria. Kelas pertama adalah faktor transkripsi, seperti faktor pernapasan nuklir11 dan 2, yang mengontrol ekspresi gen pengkode subunit dari lima kompleks pernapasan, komponen translasi mitokondria, dan enzim biosintetik heme yang dilokalisasi ke matriks mitokondria [88]. Piantadosi dkk. [89] telah menunjukkan bahwa peningkatan regulasi transkripsi yang bergantung pada Nrf2 dari faktor pernapasan nuklir 1 mendorong biogenesis mitokondria dan melindungi terhadap sitotoksisitas agen kemoterapi antrasiklin kardiotoksik doxorubicin. Sebaliknya, Zhang et al. [82] telah melaporkan bahwa aktivasi genetik Nrf2 tidak mempengaruhi ekspresi mRNA basal faktor pernapasan inti 1 di hati murine.
Kelas kedua regulator transkripsi nuklir dengan fungsi kritis dalam biogenesis mitokondria adalah koaktivator transkripsi, seperti reseptor yang diaktifkan proliferator peroksisom? coactivators (PGC) 1? dan 1 ?, yang berinteraksi dengan faktor transkripsi, mesin transkripsi basal dan penyambungan RNA, dan enzim pemodifikasi histon [88]. Ekspresi koaktivator keluarga PGC90 dipengaruhi oleh berbagai sinyal lingkungan. Pengobatan fibroblas manusia dengan sulforaphane aktivator Nrf91 menyebabkan peningkatan massa mitokondria dan induksi PGC1? dan PGC2? [1], meskipun ketergantungan potensial pada Nrf1 tidak diteliti dalam penelitian ini. Namun, tikus diabetes di mana Nrf92 diaktifkan oleh knockdown hipomorfik gen Keap2 (db / db: Keap2flox / ?: Nrf1 + / +) atau terganggu (db / db: Keap1flox / ?: Nrf2? /?) Memiliki PGC1 hati yang lebih rendah? tingkat ekspresi dari hewan kontrol (db / db: Keap2flox / +: Nrf1 + / +) [1]. Tidak ada perbedaan tingkat mRNA untuk PGC2? terlihat pada hati mencit nondiabetes yang merupakan WT atau Nrf93-KO, sedangkan level ini lebih rendah pada hewan pengekspresian berlebih Nrf1 (Keap2-KD dan Keap2-KO khusus hati) [1]. Khususnya, puasa 1 jam meningkatkan level PGC82? mRNA dalam hati mencit dari semua genotipe, tetapi peningkatan secara signifikan lebih besar pada hati Nrf24-KO dibandingkan dengan tikus yang mengekspresikan WT atau Nrf1. Dibandingkan dengan WT, tikus Nrf2-KO yang mengalami infeksi septik atau cedera paru akut akibat infeksi menunjukkan regulasi transkripsi yang dilemahkan dari faktor pernapasan nukleus 2 dan PGC2? [1], [1]. Bersama-sama, pengamatan ini menunjukkan bahwa peran Nrf94 dalam mempertahankan tingkat faktor pernapasan inti 95 dan PGC2? rumit dan menjadi paling menonjol dalam kondisi stres.
Selain ekspresi gen yang mengkode protein mitokondria, biogenesis mitokondria membutuhkan sintesis nukleotida. Aktivasi genetik Nrf2 meningkatkan biosintesis purin dengan mengatur jalur pentosa fosfat dan metabolisme folat dan glutamin, terutama pada sel yang berkembang biak dengan cepat (Gbr. 2) [24]. Analisis transkriptom mutan Drosophila kekurangan untuk mitokondria serin / treonin protein kinase PTEN-induced putative kinase 1 (PINK1) telah menunjukkan bahwa disfungsi mitokondria menyebabkan peningkatan regulasi transkripsi gen yang mempengaruhi metabolisme nukleotida [96], menunjukkan bahwa biosintesis nukleotida ditingkatkan mewakili mekanisme perlindungan terhadap konsekuensi neurotoksik dari defisiensi PINK1. Nrf2 mengatur ekspresi fosforibosil pirofosfat amidotransferase (PPAT), yang mengkatalisis masuk ke jalur biosintetik nukleotida purin de novo, dan mitokondria methylenetetrahydrofolate dehydrogenase 2 (MTHFD2) (Gbr. 2). Yang terakhir adalah enzim bifungsional dengan aktivitas dehidrogenase dan siklohidrolase yang sangat penting dalam menyediakan glisin dan format sebagai sumber unit satu karbon untuk biosintesis purin dalam sel yang tumbuh dengan cepat [97]. Oleh karena itu, aktivasi Nrf2 mungkin bersifat protektif dan dapat membalikkan disfungsi mitokondria pada defisiensi PINK1. Memang, aktivasi farmakologis Nrf2 oleh sulforaphane, atau triterpenoid RTA-408, mengembalikan m dan melindungi sel-sel yang kekurangan PINK1 terhadap toksisitas dopamin [98]. Meskipun mekanisme yang mendasari tampak kompleks, bersama-sama, temuan ini menunjukkan bahwa aktivitas Nrf2 dapat mempengaruhi biogenesis mitokondria dengan mempengaruhi tingkat ekspresi faktor transkripsi kritis dan koaktivator, serta dengan meningkatkan biosintesis nukleotida.
Nrf2 dan Integritas Mitokondria
Meskipun bukti langsung tidak selalu tersedia, ada indikasi kuat bahwa Nrf2 penting untuk integritas mitokondria, terutama di bawah kondisi stres oksidatif. Mitokondria yang diisolasi dari otak dan hati tikus yang telah diberikan satu dosis Nrf2 aktivator sulforaphane resisten terhadap pembukaan pori permeabilitas transisi mitokondria (mPTP) yang disebabkan oleh oksidasi tert-butilhidroperoksida [99], [100]. The mPTP, sebuah kompleks yang memungkinkan membran dalam mitokondria menjadi permeabel untuk molekul dengan massa hingga 1500 Da, baru-baru ini diidentifikasi akan terbentuk dari dimer dari F0F1-ATP synthase [101]. Resistensi yang diperantarai sulforaphane terhadap pembukaan mPTP berkorelasi dengan peningkatan pertahanan antioksidan, dan kadar mitokondria GSH, glutathione peroxidase 1, enzim malat 3, dan thioredoxin 2 semuanya diregulasi dalam fraksi mitokondria yang diisolasi dari hewan yang diberi sulforaphane [100].
Kerusakan protein mitokondria dan gangguan respirasi yang disebabkan oleh produk peroksidasi lipid elektrofilik 4-hidroksi-2-nonenal dilemahkan di mitokondria yang diisolasi dari korteks serebral tikus yang diobati dengan sulforaphane [102]. Dalam sel epitel ginjal tikus dan ginjal, sulforaphane melindungi terhadap toksisitas cisplatin- dan gentamisin yang diinduksi dan hilangnya m [103], [104]. Perlindungan terhadap panel oksidan (superoksida, hidrogen peroksida, peroksinitrit) dan elektrofil (4-hidroksi-2-nonenal dan akrolein) dan peningkatan pertahanan antioksidan mitokondria juga telah diamati pada pengobatan sel otot polos aorta tikus dengan sulforaphane [105 ]. Dalam model cedera ginjal akut yang diinduksi kontras, prasyarat iskemik ekstremitas baru-baru ini terbukti memiliki efek perlindungan, termasuk penghambatan pembukaan mPTP dan pembengkakan mitokondria, dengan aktivasi Nrf2 sebagai akibat dari penghambatan GSK3? [106].
Mitofagi, proses dimana mitokondria disfungsional secara selektif ditelan oleh autofagosom dan dikirim ke lisosom untuk didegradasi dan didaur ulang oleh sel, sangat penting untuk homeostasis mitokondria [107], [108]. Meskipun tidak ada hubungan kausatif antara Nrf2 dan mitofagi yang telah ditetapkan, terdapat bukti bahwa faktor transkripsi mungkin penting dalam pengendalian kualitas mitokondria dengan berperan dalam mitofagi. Ini mungkin sangat menonjol dalam kondisi stres oksidatif. Jadi, dalam model sepsis, peningkatan kadar penanda autofagosom rantai ringan MAP1 3-II (LC3-II) dan protein kargo p62 pada 24 jam pasca infeksi ditekan pada Nrf2-KO dibandingkan dengan tikus WT [109] . Sebuah penginduksi molekul kecil dari mitofagi (disebut penginduksi mitofagi yang dimediasi p62, PMI) baru-baru ini ditemukan; senyawa 1,4-difenil-1,2,3-triazol ini awalnya dirancang sebagai aktivator Nrf2 yang mengganggu interaksi faktor transkripsi dengan Keap1 [110]. Mirip dengan sel di mana Nrf2 secara genetik diregulasi (Keap1-KD atau Keap1-KO), sel yang terpapar PMI memiliki istirahat yang lebih tinggi. Yang penting, peningkatan lokalisasi LC3 mitokondria yang diamati setelah pengobatan PMI pada sel WT tidak terjadi pada sel Nrf2-KO, menunjukkan keterlibatan Nrf2.
Terakhir, analisis ultrastruktural pada bagian hati telah menunjukkan adanya mitokondria yang membengkak dengan mengurangi crista dan membran yang terganggu pada hepatosit Nrf2-KO, tetapi bukan WT, tikus yang diberi diet tinggi lemak selama 24 minggu; terutama, hati ini menunjukkan bukti yang jelas dari stres oksidatif dan peradangan [68]. Dapat disimpulkan bahwa Nrf2 memiliki peran penting dalam menjaga integritas mitokondria dalam kondisi stres oksidatif dan inflamasi.
Sulforaphane dan Dampaknya pada Kanker, Kematian, Penuaan, Otak dan Perilaku, Penyakit Jantung & Lainnya
Isothiocyanate adalah beberapa senyawa tanaman yang paling penting yang bisa Anda dapatkan dalam diet Anda. Dalam video ini saya membuat kasus yang paling komprehensif untuk mereka yang pernah dibuat. Rentang perhatian yang pendek? Lewati ke topik favorit Anda dengan mengklik salah satu poin waktu di bawah ini. Garis waktu penuh di bawah ini.
Bagian utama:
00: 01: 14 - Kanker dan kematian
00: 19: 04 - Penuaan
00: 26: 30 - Otak dan perilaku
00: 38: 06 - Rekap terakhir
00: 40: 27 - Dosis
Lini waktu penuh:
00: 00: 34 - Pengantar sulforaphane, fokus utama dari video.
00: 01: 14 - konsumsi sayuran Cruciferous dan pengurangan dalam semua penyebab kematian.
00: 02: 12 - Risiko kanker prostat.
00: 02: 23 - Risiko kanker kandung kemih.
00: 02: 34 - Kanker paru-paru berisiko pada perokok.
00: 02: 48 - Risiko kanker payudara.
00: 03: 13 - Hipotetis: bagaimana jika Anda sudah menderita kanker? (intervensi)
00: 03: 35 - Mekanisme yang masuk akal mengemudi kanker dan data asosiatif kematian.
00: 04: 38 - Sulforaphane dan kanker.
00: 05: 32 - Hewan bukti yang menunjukkan efek kuat dari ekstrak kecambah brokoli pada perkembangan tumor kandung kemih pada tikus.
00: 06: 06 - Pengaruh suplementasi langsung sulforaphane pada pasien kanker prostat.
00: 07: 09 - Bioakumulasi metabolit isothiocyanate dalam jaringan payudara yang sebenarnya.
00: 08: 32 - Penghambatan sel induk kanker payudara.
00: 08: 53 - Pelajaran sejarah: brassica ditetapkan memiliki sifat-sifat kesehatan bahkan di Roma kuno.
00: 09: 16 - Kemampuan Sulforaphane untuk meningkatkan ekskresi karsinogen (benzena, akrolein).
00: 09: 51 - NRF2 sebagai saklar genetik melalui elemen respons antioksidan.
00: 10: 10 - Bagaimana aktivasi NRF2 meningkatkan ekskresi karsinogen melalui glutathione-S-conjugates.
00: 10: 34 - kubis Brussel meningkatkan glutathione-S-transferase dan mengurangi kerusakan DNA.
00: 11: 20 - Broccoli sprout drink meningkatkan ekskresi benzena oleh 61%.
00: 13: 31 - Broccoli sprout homogenate meningkatkan enzim antioksidan di saluran napas bagian atas.
00: 15: 45 - konsumsi sayuran Cruciferous dan kematian penyakit jantung.
00: 16: 55 - Bubuk tunas brokoli meningkatkan lipid darah dan risiko penyakit jantung secara keseluruhan pada penderita diabetes tipe 2.
00: 19: 04 - Awal dari bagian penuaan.
00: 19: 21 - Diet yang diperkaya Sulforaphane meningkatkan masa hidup kumbang dari 15 ke 30% (dalam kondisi tertentu).
00: 20: 34 - Pentingnya peradangan rendah untuk umur panjang.
00: 22: 05 - Sayuran dan tunas kecambah brokoli tampaknya mengurangi beragam penanda inflamasi pada manusia.
00: 23: 40 - Rekap video pertengahan: kanker, bagian penuaan
00: 24: 14 - Studi pada tikus menunjukkan sulforaphane dapat meningkatkan fungsi imun adaptif di usia tua.
00: 25: 18 - Sulforaphane meningkatkan pertumbuhan rambut pada model tikus botak. Gambar di 00: 26: 10.
00: 26: 30 - Awal dari bagian otak dan perilaku.
00: 27: 18 - Pengaruh ekstrak kecambah brokoli pada autisme.
00: 27: 48 - Pengaruh glucoraphanin pada skizofrenia.
00: 28: 17 - Mulai dari diskusi depresi (mekanisme dan studi yang masuk akal).
00: 31: 21 - Studi mouse menggunakan 10 model yang berbeda dari depresi yang diinduksi stres menunjukkan sulforaphane sama efektifnya dengan fluoxetine (prozac).
00: 32: 00 - Studi menunjukkan konsumsi langsung glukoraphanin pada tikus juga efektif mencegah depresi dari model stres kekalahan sosial.
00: 33: 01 - Awal dari bagian neurodegenerasi.
00: 33: 30 - Sulforaphane dan penyakit Alzheimer.
00: 33: 44 - Sulforaphane dan penyakit Parkinson.
00: 33: 51 - Sulforaphane dan penyakit Hungtington.
00: 34: 13 - Sulforaphane meningkatkan protein heat shock.
00: 34: 43 - Awal dari bagian cedera otak traumatis.
00: 35: 01 - Sulforaphane disuntikkan segera setelah TBI meningkatkan daya ingat (studi pada tikus).
00: 35: 55 - Sulforaphane dan plastisitas neuronal.
00: 36: 32 - Sulforaphane meningkatkan pembelajaran dalam model diabetes tipe II pada tikus.
00: 37: 19 - Sulforaphane dan duchenne muscular dystrophy.
00: 37: 44 - Myostatin inhibition dalam sel-sel satelit otot (in vitro).
00: 38: 06 - Rekap video-ulang: mortalitas dan kanker, kerusakan DNA, stres oksidatif dan peradangan, ekskresi benzena, penyakit kardiovaskular, diabetes tipe II, efek pada otak (depresi, autisme, skizofrenia, neurodegenerasi), jalur NRF2.
00: 40: 27 - Pikiran tentang mencari tahu kecambah brokoli atau sulforaphane.
00: 41: 01 - Anekdot saat bertumbuh di rumah.
00: 43: 14 - Pada suhu memasak dan aktivitas sulforaphane.
00: 43: 45 - Konversi bakteri usus dari sulforaphane dari glucoraphanin.
00: 44: 24 - Suplemen bekerja lebih baik ketika dikombinasikan dengan myrosinase aktif dari sayuran.
00: 44: 56 - Teknik memasak dan sayuran silangan.
00: 46: 06 - Isothiocyanate sebagai goitrogens.
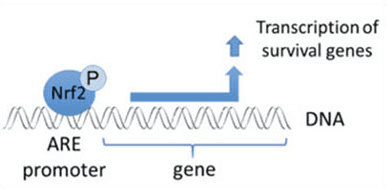
Nrf2 adalah faktor transkripsi yang memainkan peran penting dalam sistem pertahanan antioksidan seluler tubuh manusia. Unsur responsif antioksidan, atau ARE, adalah mekanisme pengaturan gen. Banyak penelitian telah menunjukkan bahwa Nrf2, atau NF-E2-terkait faktor 2, mengatur berbagai macam gen yang digerakkan oleh ARE di beberapa jenis sel. Nrf2 juga ditemukan memainkan peran penting dalam perlindungan seluler dan anti-karsinogenisitas, yang menunjukkan bahwa Nrf2 mungkin merupakan pengobatan yang efektif dalam pengelolaan penyakit neurodegeneratif dan kanker yang diyakini disebabkan oleh stres oksidatif. Dr Alex Jimenez DC, CCST Insight
Catatan Penutup
Meskipun banyak pertanyaan masih tetap terbuka, bukti eksperimental yang tersedia jelas menunjukkan bahwa Nrf2 adalah pemain penting dalam pemeliharaan homeostasis mitokondria dan integritas struktural. Peran ini menjadi sangat penting dalam kondisi stres oksidatif, elektrofilik, dan inflamasi ketika kemampuan untuk meningkatkan respons sitoprotektif yang dimediasi Nrf2 mempengaruhi kesehatan dan kelangsungan hidup sel dan organisme secara keseluruhan. Peran Nrf2 dalam fungsi mitokondria merupakan lapisan lain dari mekanisme sitoprotektif luas yang diatur oleh faktor transkripsi ini. Karena banyak kondisi patologis manusia yang memiliki stres oksidatif, peradangan, dan disfungsi mitokondria sebagai komponen penting dari patogenesis mereka, aktivasi farmakologi Nrf2 menjanjikan untuk pencegahan penyakit dan pengobatan. Pemahaman yang komprehensif tentang mekanisme yang tepat dimana Nrf2 mempengaruhi fungsi mitokondria sangat penting untuk desain rasional dari uji klinis masa depan dan dapat menawarkan biomarker baru untuk memantau keampuhan terapeutik.
Tujuan artikel di atas adalah untuk membahas - sekaligus menunjukkan - peran yang muncul dari Nrf2 dalam fungsi mitokondria. Nrf2, atau faktor faktor 2 terkait eritroid nuklir, adalah pengatur resistensi seluler yang muncul terhadap oksidan yang dapat berkontribusi pada stres oksidatif, memengaruhi fungsi seluler dan mengarah pada perkembangan toksisitas, penyakit kronis, dan bahkan kanker. Sementara produksi oksidan dalam tubuh manusia dapat melayani berbagai tujuan, termasuk pembelahan sel, peradangan, fungsi kekebalan tubuh, autofagi, dan respons stres, penting untuk mengontrol produksi berlebih mereka untuk mencegah masalah kesehatan. Cakupan informasi kami terbatas pada masalah chiropraktik dan kesehatan tulang belakang. Untuk membahas pokok bahasan ini, jangan ragu untuk bertanya kepada Dr. Jimenez atau hubungi kami di 915-850-0900 .
Nyeri punggung adalah salah satu penyebab paling umum dari kecacatan dan hari-hari yang terlewat di tempat kerja di seluruh dunia. Nyeri punggung dikaitkan dengan alasan paling umum kedua untuk kunjungan kantor dokter, kalah jumlah hanya oleh infeksi saluran pernapasan atas. Sekitar 80 persen populasi akan mengalami sakit punggung setidaknya sekali sepanjang hidup mereka. Tulang belakang adalah struktur kompleks yang terdiri dari tulang, persendian, ligamen, dan otot, di antara jaringan lunak lainnya. Karena itu, cedera dan / atau kondisi yang semakin parah, seperti cakram hernia, pada akhirnya dapat menimbulkan gejala nyeri punggung. Cedera olahraga atau cedera kecelakaan mobil sering kali menjadi penyebab paling sering dari nyeri punggung, namun terkadang gerakan yang paling sederhana bisa menimbulkan rasa sakit. Untungnya, pilihan pengobatan alternatif, seperti perawatan chiropractic, dapat membantu meringankan sakit punggung melalui penggunaan penyesuaian tulang belakang dan manipulasi manual, yang pada akhirnya meningkatkan pereda nyeri. �
Nrf2 mendukung aktivasi sekelompok antioksidan dan enzim dan gen detoksifikasi yang melindungi tubuh manusia dari efek masalah kesehatan yang terkait dengan peningkatan tingkat stres oksidatif, seperti penyakit Alzheimer. Berbagai zat alami telah ditunjukkan untuk mengaktifkan jalur Nrf2, yang dapat membantu mengelola gejala penyakit neurodegeneratif. Tujuan artikel di bawah ini adalah untuk membahas peran penting Nrf2 yang disebabkan oleh peradangan kronis.
Abstrak
Peradangan adalah ciri paling umum dari banyak penyakit kronis dan komplikasi, sementara memainkan peran penting dalam karsinogenesis. Beberapa penelitian telah menunjukkan bahwa Nrf2 berkontribusi pada proses anti-inflamasi dengan mengatur perekrutan sel inflamasi dan mengatur ekspresi gen melalui elemen respon antioksidan (ARE). Jalur pensinyalan Keap1 (Kelch-like ECH-related protein) / Nrf2 (NF-E2 p45-related factor 2) / ARE terutama mengatur ekspresi gen anti-inflamasi dan menghambat perkembangan peradangan. Oleh karena itu, identifikasi fitokimia antiinflamasi baru yang bergantung pada Nrf2 telah menjadi titik kunci dalam penemuan obat. Dalam ulasan ini, kami membahas anggota jalur sinyal Keap1 / Nrf2 / ARE dan gen hilirnya, efek jalur ini pada model hewan penyakit inflamasi, dan crosstalk dengan jalur NF-? B. Selain itu kami juga membahas tentang regulasi NLRP3 inflammasome oleh Nrf2. Selain itu, kami merangkum skenario terkini dari pengembangan fitokimia anti-inflamasi dan lainnya yang memediasi jalur pensinyalan Nrf2 / ARE.
Kata kunci: Nrf2, Keap1, ARE, Peradangan, Stres oksidatif, Fitokimia
Peradangan adalah proses kompleks yang terjadi ketika jaringan terinfeksi atau terluka oleh rangsangan berbahaya seperti patogen, kerusakan, atau iritasi. Sel imun, pembuluh darah, dan mediator molekuler terlibat dalam respons perlindungan ini [1]. Peradangan juga merupakan fenomena patologis yang terkait dengan berbagai kondisi penyakit yang disebabkan terutama oleh faktor fisik, kimia, biologis, dan psikologis. Tujuan peradangan adalah untuk membatasi dan menghilangkan penyebab kerusakan sel, membersihkan dan / atau menyerap sel dan jaringan nekrotik, dan memulai perbaikan jaringan. Dua bentuk peradangan yang berbeda dibedakan: akut dan kronis. Peradangan akut membatasi diri dan bermanfaat bagi tuan rumah, tetapi peradangan kronis yang berkepanjangan adalah fitur umum dari banyak penyakit kronis dan komplikasi. Infiltrasi langsung oleh banyak sel kekebalan mononuklear seperti monosit, makrofag, limfosit, dan sel plasma, serta produksi sitokin inflamasi, menyebabkan peradangan kronis. Diketahui bahwa peradangan kronis memainkan peran penting dalam karsinogenesis [2]. Secara umum, jalur sinyal pro dan anti-inflamasi berinteraksi dalam proses inflamasi normal.
Dalam proses inflamasi patologis, sel mast, monosit, makrofag, limfosit, dan sel kekebalan lainnya pertama kali diaktifkan. Kemudian sel-sel tersebut direkrut ke lokasi luka, menghasilkan generasi spesies oksigen reaktif (ROS) yang merusak makromolekul termasuk DNA. Pada saat yang sama, sel inflamasi ini juga menghasilkan mediator inflamasi dalam jumlah besar seperti sitokin, kemokin, dan prostaglandin. Mediator ini selanjutnya merekrut makrofag ke lokasi inflamasi yang terlokalisasi dan secara langsung mengaktifkan beberapa kaskade transduksi sinyal dan faktor transkripsi yang terkait dengan inflamasi. Jalur pensinyalan NF-? B (faktor inti kappa B), MAPK (mitogen-activated protein kinase), dan JAK (janus kinase) -STAT (transduser sinyal dan penggerak transkripsi) jalur pensinyalan terlibat dalam pengembangan jalur klasik inflamasi [3], [4], [5]. Penelitian sebelumnya telah mengungkapkan bahwa faktor transkripsi Nrf2 (NF-E2 p45-related factor 2) mengatur ekspresi enzim detoksifikasi fase II termasuk NADPH, NAD (P) H kuinon oksidoreduktase 1, glutathione peroksidase, ferritin, heme oxygenase-1 (HO -1), dan gen antioksidan yang melindungi sel dari berbagai cedera melalui efek anti-inflamasi mereka, sehingga mempengaruhi perjalanan penyakit [6], [7], [8].
Mempertimbangkan temuan yang luar biasa ini, pengembangan obat terapeutik yang ditargetkan untuk penyakit radang melalui jalur sinyal telah menarik banyak perhatian dalam beberapa tahun terakhir. Dalam ulasan ini, kami meringkas penelitian tentang Keap1 (protein terkait ECH Kelch-like) / Nrf2 (NF-E2 p45-terkait faktor 2) / ARE (elemen respons antioksidan) jalur pensinyalan pada peradangan.
Struktur dan Pengaturan Nrf2
Keap1-Dependent Peraturan Nrf2
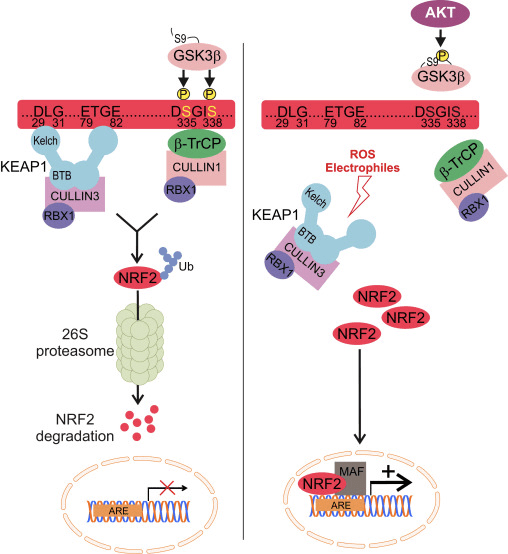
Nrf2 termasuk dalam subfamili Cap n Collar (CNC) dan terdiri dari tujuh domain fungsional, Neh (homologi Nrf2-ECH) 1 hingga Neh7 [9], [10]. Neh1 adalah domain CNC-bZIP yang memungkinkan Nrf2 melakukan heterodimerisasi dengan protein muskuloaponeurotik fibrosarkoma (Maf) kecil, DNA, dan mitra transkripsi lainnya serta membentuk kompleks inti dengan enzim konjugasi ubiquitin UbcM2 [11], [12]. Neh2 mengandung dua motif penting yang dikenal sebagai DLG dan ETGE, yang penting untuk interaksi antara Nrf2 dan regulator negatif Keap1 [13], [14].
Keap1 adalah adaptor substrat untuk E3 ubiquitin ligase berbasis cullin, yang menghambat aktivitas transkripsi Nrf2 melalui ubiquitination dan degradasi proteasomal dalam kondisi normal [15], [16], [17]. Domain KELCH dari homodimer Keap1 mengikat dengan motif DLG dan ETGE dari domain Nrf2-Neh2 di sitosol, di mana ETGE bertindak sebagai engsel dengan afinitas dan DLG yang lebih tinggi bertindak sebagai gerendel [18]. Di bawah tekanan oksidatif atau setelah terpapar pada aktivator Nrf2, Nrf2 berdisosiasi dari ikatan Keap1 karena modifikasi tiol residu cysteine Keap1 yang pada akhirnya mencegah Nrf2 ubiquitination dan degradasi proteasom [19]. Kemudian Nrf2 mentranslokasi ke dalam nukleus, heterodimerizes dengan protein Maf kecil, dan melakukan transactivates baterai baterai ARE (Gambar 1A). Terminal karboksi Neh3 bertindak sebagai domain transactivation dengan berinteraksi dengan aktivator transkripsi yang dikenal sebagai CHD6 (protein pengikat DNA chromo-ATPase / helicase) [20]. Neh4 dan Neh5 juga bertindak sebagai domain transaktivasi, tetapi mengikat ke aktivator transkripsional lain yang dikenal sebagai CBP (protein pengikat protein pengikat-respons-elemen-cAMP) [21]. Selain itu, Neh4 dan Neh5 berinteraksi dengan kofaktor nuklir RAC3 / AIB1 / SRC-3, yang mengarah ke ekspresi gen ARE yang ditargetkan Nrf2 yang ditargetkan [22]. Neh5 memiliki sinyal ekspor nuklir redoks-sensitif yang sangat penting untuk regulasi dan lokalisasi seluler Nrf2 [23].
Gambar 1 Keap1-dependent dan regulasi independen Nrf2. (A) Di bawah kondisi basal, Nrf2 diasingkan dengan Keap1 oleh dua motifnya (ETGE dan DLG) yang mengarah ke ubiquitination yang dimediasi CUL3 diikuti oleh degradasi proteasome. Di bawah tekanan oksidatif, Nrf2 berdisosiasi dari Keap1, berpindah ke nukleus dan mengaktifkan baterai gen ARE. (B) GSK3 memfosforilasi Nrf2 dan ini memfasilitasi pengenalan Nrf2 oleh? -TrCP untuk ubiquitination yang dimediasi CUL1 dan degradasi proteasom berikutnya. (C) p62 diasingkan dengan Keap1, menyebabkan degradasi autofagiknya, pembebasan Nrf2, dan peningkatan pensinyalan Nrf2.
Peraturan Nrf1 Independen Keap2
Bukti yang muncul telah mengungkapkan mekanisme baru regulasi Nrf2 yang tidak bergantung pada Keap1. Domain Neh6 yang kaya serin dari Nrf2 memainkan peran penting dalam regulasi ini dengan mengikat dua motifnya (DSGIS dan DSAPGS) ke? -Transdusin protein yang mengandung berulang (? -TrCP) [24]. ? -TrCP adalah reseptor substrat untuk kompleks ligase ubiquitin Skp1 Cul1 Rbx1 / Roc1 yang menargetkan Nrf2 untuk ubiquitination dan degradasi proteasomal. Glikogen sintase kinase-3 adalah protein penting yang terlibat dalam stabilisasi dan regulasi Nrf1 independen-Keap2; itu memfosforilasi Nrf2 di domain Neh6 untuk memfasilitasi pengenalan Nrf2 oleh? -TrCP dan degradasi protein selanjutnya [25] (Gbr. 1B).
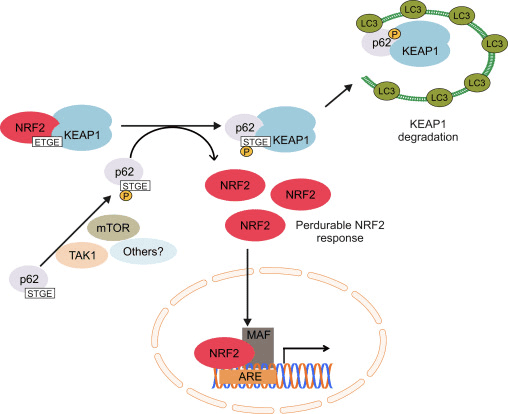
Regulator Nrf2 lainnya
Garis bukti lain telah menunjukkan jalur non-kanonik aktivasi Nrf62 yang bergantung pada p2 di mana p62 mengasingkan Keap1 ke degradasi autophagic yang pada akhirnya mengarah pada stabilisasi Nrf2 dan transaktivasi gen tergantung Nrf2 [26], [27], [ 28], [29] (Gbr. 1C).
Bukti yang terkumpul menunjukkan bahwa beberapa miRNA memainkan peran penting dalam regulasi aktivitas Nrf2 [30]. Sangokoya dkk. [31] menunjukkan bahwa miR-144 secara langsung menurunkan aktivitas Nrf2 dalam garis sel limfoblas K562, sel progenitor eritroid primer manusia, dan retikulosit penyakit sel sabit. Studi menarik lainnya pada sel epitel payudara manusia menunjukkan bahwa miR-28 menghambat Nrf2 melalui mekanisme independen Keap1 [32]. Demikian pula, miRNA seperti miR-153, miR-27a, miR-142-5p, dan miR144 menurunkan regulasi ekspresi Nrf2 dalam garis sel SH-SY5Y neuronal [33]. Singh dkk. [34] menunjukkan bahwa ekspresi ektopik miR-93 menurunkan ekspresi gen yang diatur Nrf2 dalam model tikus karsinogenesis mammae yang diinduksi 17? -Estradiol (E2).
Penemuan baru-baru ini dari laboratorium kami mengidentifikasi penghambat endogen Nrf2 yang dikenal sebagai reseptor alfa retinoik X (RXR?). RXR? adalah reseptor inti, berinteraksi dengan domain Neh7 dari Nrf2 (residu asam amino 209 316) melalui domain pengikat DNA (DBD), dan secara khusus menghambat aktivitas Nrf2 di dalam nukleus. Selain itu, reseptor nuklir lain seperti reseptor yang diaktifkan proliferator peroksisom- ?, ER ?, reseptor yang berhubungan dengan estrogen- ?, dan reseptor glukokortikoid juga telah dilaporkan menjadi penghambat endogen aktivitas Nrf2 [9], [10].
Peran Anti-inflamasi Nrf2 / HO-1 Axis
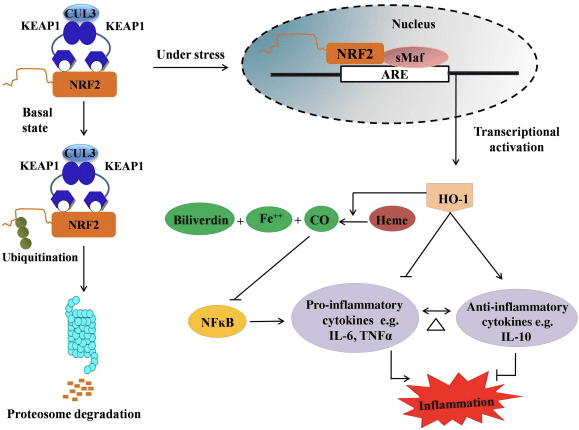
HO-1 adalah isoform yang dapat diinduksi dan enzim pembatas laju yang mengkatalisis degradasi heme menjadi karbon monoksida (CO) dan besi bebas, dan biliverdin menjadi bilirubin. Degradasi enzimatik bebas pro-inflamasi heme serta produksi senyawa anti-inflamasi seperti CO dan bilirubin memainkan peran utama dalam menjaga efek perlindungan dari HO-1 (Gambar 2).
Gambar 2 Ikhtisar jalur Nrf2 / HO-1. Di bawah kondisi basal, Nrf2 mengikat repressor Keap1 yang mengarah ke ubiquitination diikuti oleh degradasi proteasome. Selama stres oksidatif, Nrf2 bebas berpindah ke nukleus, di mana ia dimerisasi dengan anggota keluarga Maf kecil dan berikatan dengan gen ARE seperti HO-1. HO-1 yang diatur secara berlebihan mengkatalisis heme menjadi CO, bilirubin, dan besi bebas. CO bertindak sebagai penghambat jalur NF-? B yang menyebabkan penurunan ekspresi sitokin proinflamasi, sedangkan bilirubin juga bertindak sebagai antioksidan. Lebih lanjut, HO-1 secara langsung menghambat sitokin proinflamasi serta mengaktifkan sitokin anti inflamasi, sehingga mengarah pada keseimbangan proses inflamasi.
Nrf2 menginduksi gen HO-1 dengan meningkatkan mRNA dan ekspresi protein dan merupakan salah satu gen klasik Nrf2 yang diatur yang banyak digunakan dalam berbagai penelitian in vitro dan in vivo. Beberapa penelitian telah menunjukkan bahwa HO-1 dan metabolitnya memiliki efek antiinflamasi yang signifikan yang dimediasi oleh Nrf2. Peningkatan ekspresi HO-1 yang dimediasi oleh Nrf2 teraktivasi menyebabkan penghambatan pensinyalan NF-B menghasilkan penurunan cedera mukosa usus dan disfungsi taut-ketat pada model transplantasi hati tikus Sprague-Dawley jantan [35]. Peningkatan regulasi ekspresi HO-2 yang bergantung pada Nrf1 dapat melindungi myoblas C2C12 yang diturunkan dari tikus dari sitotoksisitas H2O2 [36]. HO-2 yang bergantung pada Nrf1 berdampak pada respons inflamasi yang dimediasi lipopolisakarida (LPS) di RAW264.7- atau makrofag sel busa yang diturunkan dari makrofag peritoneal tikus. Aktivitas Nrf2 desensitized foam cell makrofag fenotipe dan mencegah peradangan makrofag yang tidak moderat, yang memainkan peran penting dalam perkembangan aterosklerosis [37]. Sumbu Nrf2 / HO-1 mempengaruhi sel mikroglial BV2 tikus yang diinduksi LPS dan sel HT22 hipokampus tikus, yang berdampak pada peradangan saraf. Peningkatan regulasi ekspresi HO-1 melalui jalur Nrf2 pada sel mikroglial BV2 tikus yang mempertahankan kematian sel sel HT22 hipokampus tikus [38]. Lebih lanjut, molekul hibrida berbasis kobalt (HYCO) yang menggabungkan penginduksi Nrf2 dengan pelepas karbon monoksida (CO) meningkatkan ekspresi Nrf2 / HO-1, membebaskan CO dan menggunakan aktivitas anti-inflamasi secara in vitro. HYCOs juga mengatur jaringan HO-1 dan mengantarkan CO dalam darah setelah pemberian in vivo, mendukung potensi penggunaannya melawan kondisi inflamasi [39]. Peningkatan regulasi Nrf2 / HO-1 mengurangi inflamasi dengan meningkatkan aktivitas eferositik makrofag murine yang diobati dengan taurine chloramines [40]. Secara keseluruhan, model eksperimental yang dijelaskan di atas mengungkapkan bahwa sumbu Nrf2 / HO-1 memainkan peran utama dalam fungsi anti-inflamasi, menunjukkan bahwa Nrf2 adalah target terapeutik pada penyakit terkait peradangan.
Selain itu, produk sampingan dari HO-1 seperti CO, bilirubin, bertindak sebagai antioksidan kuat selama stres oksidatif dan kerusakan sel [41], [42]; itu menekan encephalomyelitis autoimmune dan hepatitis [43], [44]; dan melindungi tikus dan tikus terhadap syok endotoxic dengan mencegah generasi iNOS dan NO [45], [46], [47]. Selain itu, Bilirubin mengurangi aktivasi dan disfungsi endotel [48]. Menariknya, bilirubin mengurangi transmigrasi leukosit endotel melalui molekul adhesi-1 [49]. Referensi khusus ini menunjukkan tidak hanya HO-1 bertindak sebagai agen anti-inflamasi yang kuat tetapi juga metabolitnya.
Mediator Inflamasi dan Enzymes dihambat oleh Nrf2
Sitokin dan Kemokin
Sitokin adalah protein dengan berat molekul rendah dan polipeptida yang disekresikan oleh berbagai sel; mereka mengatur pertumbuhan sel, diferensiasi, dan fungsi kekebalan, dan terlibat dalam peradangan dan penyembuhan luka. Sitokin termasuk interleukin (ILs), interferon, faktor nekrosis tumor (TNF), faktor perangsang koloni, kemokin, dan faktor pertumbuhan. Beberapa sitokin dihitung sebagai mediator pro-inflamasi sedangkan yang lain memiliki fungsi anti-inflamasi. Paparan hasil stres oksidatif dalam produksi berlebih sitokin yang menyebabkan stres oksidatif dalam sel target. Beberapa sitokin proinflamasi diproduksi berlebih ketika NF-? B diaktivasi oleh stres oksidatif. Selanjutnya, stres oksidatif pro-inflamasi menyebabkan aktivasi lebih lanjut dari NF-? B dan produksi sitokin yang berlebihan. Aktivasi sistem Nrf2 / ARE memainkan peran penting dalam mengganggu siklus ini. Kemokin adalah keluarga sitokin kecil, yang peran utamanya adalah memandu migrasi sel inflamasi. Mereka berfungsi terutama sebagai chemoattractant untuk leukosit, monosit, neutrofil, dan sel efektor lainnya.
Telah dilaporkan bahwa aktivasi Nrf2 mencegah peningkatan regulasi transkripsi yang diinduksi LPS dari sitokin proinflamasi, termasuk IL-6 dan IL-1? [50]. IL-1? dan produksi IL-6 juga meningkat pada Nrf2? /? tikus dengan kolitis akibat dekstran sulfat [51], [52]. Nrf2 menghambat produksi IL-17 hilir dan faktor inflamasi lainnya Th1 dan Th17, dan menekan proses penyakit dalam model eksperimental multiple sclerosis, autoimun ensefalitis [53]. Gen anti-oksidan yang bergantung pada Nrf2 HO-1, NQO-1, Gclc, dan Gclm memblokir TNF- ?, IL-6, monosit kemo atraktan protein-1 (MCP1), protein inflamasi makrofag-2 (MIP2), dan inflamasi. mediator. Tetapi dalam kasus tikus Nrf2-knockout, efek anti-inflamasi tidak terjadi [54]. Neutrofil peritoneal dari tikus Nrf2-knockout yang diobati dengan LPS memiliki tingkat sitokin (TNF-? Dan IL-6) dan kemokin (MCP1 dan MIP2) yang lebih tinggi secara signifikan daripada sel tipe liar (WT) [54]. In vitro, mentransfer gen Nrf2 ke sel otot polos aorta manusia dan kelinci menekan sekresi MCP1 [8], [55], dan ekspresi HO-2 yang bergantung pada Nrf1 menekan TNF -? - terstimulasi NF-? B dan MCP-1 sekresi dalam sel endotel vena umbilikalis manusia [56]. Temuan ini mengisyaratkan bahwa, dalam menanggapi rangsangan inflamasi, peningkatan regulasi pensinyalan Nrf2 menghambat produksi berlebih dari sitokin dan kemokin pro-inflamasi serta membatasi aktivasi NF-? B.
Molekul Adhesi Sel
Molekul adhesi sel (CAM) adalah protein yang mengikat dengan sel atau dengan matriks ekstraseluler. Terletak di permukaan sel, mereka terlibat dalam pengenalan sel, aktivasi sel, transduksi sinyal, proliferasi, dan diferensiasi. Di antara CAM, ICAM-1 dan VCAM-1 adalah anggota penting dari superfamili imunoglobulin. ICAM-1 hadir dalam konsentrasi rendah di leukosit dan membran sel endotel. Setelah stimulasi sitokin, konsentrasinya meningkat secara signifikan. ICAM-1 dapat diinduksi oleh IL-1 dan TNF dan diekspresikan oleh endotel vaskular, makrofag, dan limfosit. Ini adalah ligan untuk integrin, reseptor yang ditemukan pada leukosit. Ketika jembatan ICAM-1-integrin diaktifkan, leukosit mengikat ke sel endotel dan kemudian bermigrasi ke jaringan subendotel [57]. VCAM-1 memediasi adhesi limfosit, monosit, eosinofil, dan basofil ke endotel vaskular dan berkontribusi pada perekrutan leukosit, yang pada akhirnya menyebabkan kerusakan jaringan akibat stres oksidatif. Nrf2 menghambat aktivitas promotor VCAM-1 [58]. Gen hilir yang diatur Nrf2 HO-1 dapat mempengaruhi ekspresi E-selektin dan VCAM-1, molekul adhesi yang berhubungan dengan sel endotel [59]. Ekspresi paru dari beberapa CAM seperti CD-14, TREM1, SELE, SELP, dan VCAM-1 secara signifikan lebih tinggi di Nrf2? /? tikus daripada di Nrf2 + / + tikus [60]. Nrf2 dalam sel endotel aorta manusia menekan TNF -? - menginduksi ekspresi VCAM-1 dan mengganggu TNF -? - menginduksi adhesi sel U937 monositik [8]. Ekspresi berlebihan Nrf2 juga menghambat ekspresi gen VCAM-1 yang diinduksi TNF -? - dalam sel endotel mikrovaskuler manusia [61]. Antioksidan alami asam 3-hidroksiantranilik (HA), salah satu metabolit l-triptofan yang terbentuk secara in vivo di sepanjang jalur metabolisme yang dikenal sebagai jalur kynurenine selama peradangan atau infeksi, ditemukan untuk menginduksi ekspresi HO-1 dan merangsang Nrf2 pada pusar manusia. sel endotel vena (HUVECs). Ekspresi HO-2 yang bergantung pada Nrf1 yang diinduksi oleh HA menghambat sekresi MCP-1, ekspresi VCAM-1 dan aktivasi NF-kB yang terkait dengan cedera vaskular dan inflamasi pada aterosklerosis [56]. Turunan chalcone sintetis anti-proliferatif dan anti-inflamasi 2?, 4?, 6? -Tris (methoxymethoxy) chalcone menghambat ICAM-1, sitokin pro-inflamasi IL-1 ?, dan TNF-? ekspresi dalam jaringan kolon dari tikus yang diobati dengan asam sulfonat trinitrobenzene [62]. Peningkatan regulasi Nrf2 menghambat ekspresi ICAM-1 yang diinduksi TNF -? - dalam sel epitel pigmen retina manusia yang diobati dengan likopen [63]. Semua studi ini menunjukkan bahwa Nrf2 memainkan peran kunci dalam proses inflamasi dengan mengatur migrasi dan infiltrasi sel inflamasi ke jaringan yang meradang.
Matrix Metalloproteinase (MMPs)
MMP terdapat secara luas dalam matriks ekstraseluler dan terlibat dalam proses fisiologis dan patologis seperti proliferasi sel, migrasi, diferensiasi, penyembuhan luka, angiogenesis, apoptosis, dan metastasis tumor. Telah dilaporkan bahwa sumbu Nrf2 / HO-1 menghambat MMP-9 di makrofag dan MMP-7 di sel epitel usus manusia, dan ini bermanfaat dalam pengobatan penyakit radang usus [62]. Kerusakan kulit akibat iradiasi UV lebih parah pada Nrf64-knockout dibandingkan pada tikus WT dan tingkat MMP-2 secara signifikan lebih tinggi, menunjukkan bahwa Nrf9 mengurangi ekspresi MMP-2. Oleh karena itu, Nrf9 dianggap sebagai pelindung terhadap iradiasi UV [2]. Studi lain juga melaporkan bahwa aktivasi transkripsi MMP-65 yang diatur ke bawah dalam invasi sel tumor dan peradangan diatur melalui penghambatan jalur pensinyalan NF-kB [9]. Pada cedera tulang belakang traumatis, jalur pensinyalan NF-kB juga berperan dalam mengatur tingkat mRNA MMP-66 [9]. Oleh karena itu, pada inflamasi regulasi MMPs dipengaruhi secara langsung oleh jalur Nrf67 atau secara tidak langsung melalui jalur NF-? B yang dipengaruhi Nrf2.
Cyclooxygenase-2 (COX2) dan Induced Nitric Oxide Synthase (INOS)
Serangkaian percobaan pada tikus Nrf2-knockout telah menunjukkan peran penting dalam inflamasi dan regulasi gen pro-inflamasi seperti COX-2 dan iNOS. Untuk pertama kalinya, Khor et al. melaporkan peningkatan ekspresi sitokin pro-inflamasi seperti COX-2 dan iNOS di jaringan kolon Nrf2? /? tikus dibandingkan dengan tikus WT Nrf2 + / +, menunjukkan bahwa Nrf2 menekan aktivitas mereka [51]. Laporan lain tentang pretreatment dengan sulforaphane, salah satu aktivator Nrf2 terkenal yang ada dalam sayuran silangan, menunjukkan efek antiinflamasinya dalam menghambat ekspresi TNF- ?, IL-1 ?, COX-2, dan iNOS di kedua mRNA tersebut. dan tingkat protein di makrofag peritoneal primer dari tikus Nrf2 + / + dibandingkan dengan Nrf2? /? tikus [68]. Demikian pula, hipokampus tikus Nrf2-knockout dengan inflamasi yang diinduksi LPS juga menunjukkan ekspresi penanda inflamasi yang lebih tinggi seperti iNOS, IL-6, dan TNF-? dari tikus WT [69]. Demikian juga, tikus knockout Nrf2 hipersensitif terhadap stres oksidatif yang diinduksi oleh 1-metil-4-fenil-1,2,3,6-tetrahidropiridin serta menunjukkan peningkatan mRNA dan kadar protein penanda inflamasi seperti COX-2, iNOS , IL-6, dan TNF-? [70]. Selain itu, hati dari Nrf2? /? tikus yang ditantang dengan diet kekurangan metionin dan kolin memiliki ekspresi mRNA ~ 5 kali lipat lebih tinggi dari Cox2, dan iNOS dibandingkan tikus WT dengan diet yang sama, menunjukkan peran anti-inflamasi Nrf2 [71]. Baru-baru ini, Kim et al. menunjukkan bahwa fitokimia etil piruvat memberikan efek anti-inflamasi dan anti-oksidatif dengan mengurangi ekspresi iNOS melalui pensinyalan Nrf2 dalam sel BV2. Mereka menunjukkan bahwa etil piruvat menginduksi translokasi inti Nrf2, yang pada akhirnya menghambat interaksi antara p65 dan p300, yang menyebabkan penurunan ekspresi iNOS [72]. Selanjutnya, analog karbazol LCY-2-CHO mengaktifkan Nrf2 dan menyebabkan translokasi nukleusnya, yang mengarah pada penekanan ekspresi COX2 dan iNOS [73] pada sel otot polos vaskular aorta tikus.
Peran Paradoks Nrf2 dalam Regulasi Aktivitas NLRP3 iIflammasome�
Keluarga NLR, domain pyrin yang mengandung 3 (NLRP3) inflammasome adalah kompleks multiprotein yang berfungsi sebagai reseptor pengenalan patogen (PRR) dan mengenali berbagai mikroba, sinyal stres oksidatif seperti pola molekuler terkait patogen (PAMPs), Kerusakan- molekul pola molekul terkait (DAMPs) dan ROS [74]. Inflamasom NLRP3 yang teraktivasi memediasi pembelahan caspase-1 dan sekresi sitokin interleukin-1 pro-inflamasi? (IL-1?) Yang pada akhirnya menginduksi proses kematian sel yang dikenal sebagai pyroptosis yang melindungi inang dari berbagai patogen [75]. Namun, aktivasi menyimpang dari inflammasome dikaitkan dengan penyakit misfolding protein seperti ensefalopati spongiform menular, penyakit Alzheimer, penyakit Parkinson dan juga diabetes tipe 2 [76], kanker [77], asam urat, dan aterosklerosis [78].
Pengamatan terbaru dari kelompok Rong Hu pada asosiasi Nrf2 dengan regulasi negatif dari inflammasome mengungkapkan bahwa, Nrf2 menginduksi ekspresi NQO1 yang mengarah pada penghambatan aktivasi inflamasi NLRP3, pembelahan caspase-1 dan IL-1? generasi di makrofag. Selanjutnya, aktivator Nrf2 yang terkenal, tert-butylhydroquinone (tBHQ) secara negatif mengatur transkripsi NLRP3 dengan mengaktifkan ARE dengan cara yang bergantung pada Nrf2 [79]. Selain pengamatan di atas, kelompok yang sama juga telah mengungkapkan bahwa, dimethyl fumarate (DMF) mencegah kolitis yang diinduksi DSS melalui pengaktifan jalur pensinyalan Nrf2 yang terlibat dalam translokasi nuklear Nrf2 dan penghambatan perakitan inflamasi NLRP3 [80].
Serangkaian percobaan menggunakan senyawa alami dan sintetis juga telah mengungkapkan efek penghambatan Nrf2 pada aktivasi inflamasi NLRP3. Misalnya, pengobatan epigallocatechin-3-gallate (EGCG) pada tikus lupus nephritis telah menunjukkan penurunan aktivasi inflamasi NLRP3 ginjal yang dimediasi oleh jalur pensinyalan Nrf2 [81]. Demikian juga, citral (3,7-dimethyl-2,6-octadienal), senyawa aktif utama dalam obat herbal Cina Litsea cubeba, menghambat aktivasi inflamasi NLRP3 melalui jalur pensinyalan antioksidan Nrf2 pada model tikus Accelerated and Severe Lupus Nephritis (ASLN). [82]. Demikian pula, biochanin melindungi terhadap kerusakan hati yang diinduksi LPS / GalN dengan mengaktifkan jalur Nrf2 dan menghambat aktivasi inflamasi NLRP3 pada tikus BALB / c jantan [83]. Selain itu, mangiferin juga terbukti mengatur ekspresi Nrf2 dan HO-1 dengan cara yang bergantung pada dosis dan menghambat NLRP3 hati yang diinduksi LPS / D-GalN, ASC, caspase-1, IL-1? dan TNF-? ekspresi [84].
Meskipun regulasi negatif NLRP3 oleh Nrf2, ini juga mengaktifkan fungsi inflamasi NLRP3 dan AIM2. Haitao Wen dan rekannya menemukan itu, Nrf2? /? makrofag tikus telah menunjukkan aktivasi rusak dari NLRP3 dan AIM2 Inflammasome tetapi tidak inflamasi NLRC4 [85]. Menariknya, observasi ini menggambarkan fungsi Nrf2 yang tidak diketahui dalam konteks penyakit terkait inflamasi; oleh karena itu sangat penting untuk mempelajari lebih lanjut untuk mengungkap mekanisme di mana Nrf2 mengaktifkan fungsi inflammasome sebelum mempertimbangkannya sebagai target terapeutik.
Penindasan Transplantasi Cytokin Pro-Inflamasi oleh Nrf2
Sebuah investigasi terbaru berdasarkan hasil chromatin immunoprecipitation (ChIP) -seq dan ChIP-qPCR pada makrofag tikus mengungkapkan bahwa Nrf2 berikatan dengan daerah promotor sitokin pro-inflamasi seperti IL-6 dan IL-1? dan menghambat perekrutan RNA Pol II. Akibatnya, RNA Pol II tidak dapat memproses aktivasi transkripsi IL-6 dan IL-1? yang pada akhirnya mengarah pada penghambatan ekspresi gen. Untuk pertama kalinya, kelompok Masayuki Yamamoto mengungkapkan mekanisme baru di mana Nrf2 tidak hanya mentransaktivasi gen hilirnya melalui ARE tetapi juga menekan aktivasi transkripsi gen tertentu dengan atau tanpa ARE melalui penghambatan perekrutan RNA Pol II [50].
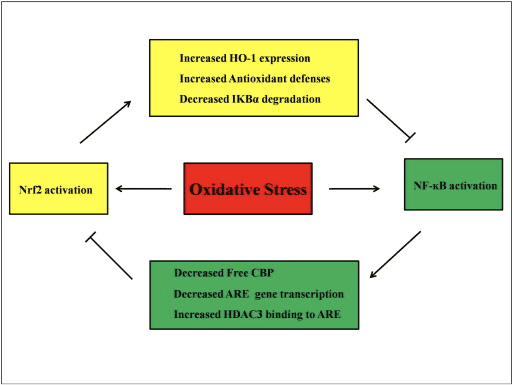
Crosstalk Antara Jalur Nrf2 dan NF-? B
NF-? B adalah protein kompleks yang bertanggung jawab untuk transkripsi DNA yang ditemukan di hampir semua jenis sel hewan dan terlibat dalam berbagai proses seperti inflamasi, apoptosis, respon imun, pertumbuhan sel, dan perkembangan. p65, protein Rel dari keluarga NF-? B, memiliki domain transaktivasi sedangkan p50 tidak dan membutuhkan heterodimerisasi dengan protein Rel untuk mengaktifkan transkripsi. Selama stres oksidatif, I? B kinase (IKK) diaktifkan dan menyebabkan fosforilasi I? B, menghasilkan pelepasan dan translokasi inti NF-? B. NF-? B menyebabkan transkripsi mediator pro-inflamasi seperti IL-6, TNF- ?, iNOS, IL-1, dan adhesi intraseluler COX-2.
Regulasi abnormal dari NF-? B telah dihubungkan dengan rheumatoid arthritis, asma, penyakit radang usus, dan gastritis akibat infeksi Helicobacter pylori [86]. Saat ini dianggap bahwa aktivitas NF-kB mempengaruhi jalur pensinyalan Keapl / Nrf2 / ARE terutama dalam tiga aspek: pertama, Keap1 menurunkan IKK? melalui ubiquitination, sehingga menghambat aktivitas NF-? B [87]. Kedua, proses inflamasi menginduksi mediator inflamasi seperti COX2 yang berasal dari siklopentenon prostaglandin 15d-PGJ2, elektrofil kuat yang bereaksi dengan Keap1 dan mengaktifkan Nrf2, sehingga memulai transkripsi gen dengan penghambatan simultan aktivitas NF-kB [58], [88] ( Gambar 3 A, B). Ketiga, NF-? B dapat digabungkan dengan co-activator transkripsi Nrf2 kompetitif CBP [89], [90] (Gbr. 3 C, D).
Gambar 3 Crosstalk antara jalur Nrf2 dan NF-? B. (A) Keap1 mengarahkan IKK ke ubiquitination yang dimediasi CUL3 dan degradasi proteasom yang pada akhirnya mengarah pada penghambatan fosforilasi NF-? B dan mekanisme ini juga bekerja sebagai pengikatan kompetitif Nrf2 dan IKK dengan Keap1. (B) Stres oksidatif mengaktifkan IKK yang memfosforilasi NF-? B, menyebabkan translokasi ke dalam nukleus dan aktivasi sitokin proinflamasi seperti COX-2. Produk terminal COX-2 yang dikenal sebagai 15d-PGJ2 bertindak sebagai penginduksi Nrf2 yang pada akhirnya mengarah pada penekanan stres oksidatif. (C) Nrf2 mengikat CBP kofaktor transkripsi bersama dengan Maf kecil dan mesin transkripsi lainnya untuk memulai ekspresi gen yang digerakkan oleh ARE. (D) Ketika NF-? B berikatan dengan CBP secara kompetitif, ia menghambat pengikatan CBP dengan Nrf2, yang mengarah pada penghambatan transaktivasi Nrf2.
Diasumsikan bahwa jalur pensinyalan Nrf2 dan NF-? B berinteraksi untuk mengontrol transkripsi atau fungsi protein target hilir. Dalam justifikasi asumsi ini banyak contoh menunjukkan bahwa aktivasi dan penghambatan langsung atau tidak langsung terjadi antara anggota jalur Nrf2 dan NF-? B (Gbr. 4). Menanggapi LPS, knockdown Nrf2 secara signifikan meningkatkan aktivitas transkripsi NF-? B dan transkripsi gen yang bergantung pada NF-? B, menunjukkan bahwa Nrf2 menghambat aktivitas NF-? B [60]. Selain itu, peningkatan ekspresi HO-2 hilir yang bergantung pada Nrf1 menghambat aktivitas NF-? B. Ketika sel kanker prostat terpapar sebentar ke? -Tochopheryl succinate, turunan dari vitamin E, ekspresi HO-1 diregulasi. Produk akhir HO-1 menghambat translokasi inti NF-? B [92]. Studi in vivo ini menunjukkan bahwa Nrf2 secara negatif mengatur jalur pensinyalan NF-kB. LPS merangsang aktivitas pengikatan DNA NF-? B dan tingkat subunit p65 dari NF-? B secara signifikan lebih tinggi pada ekstrak inti dari paru-paru Nrf2? /? daripada tikus WT, menunjukkan peran negatif Nrf2 dalam aktivasi NF-? B. Selain itu, Nrf2? /? fibroblas embrio tikus yang diobati dengan LPS dan TNF-? menunjukkan aktivasi NF-? B yang lebih menonjol yang disebabkan oleh aktivasi IKK dan I? B-? degradasi [60]. Dan pembersihan virus pernapasan syncytial menurun secara signifikan sementara aktivitas pengikatan DNA NF-? B meningkat di Nrf2? /? tikus dibandingkan dengan tikus WT [93]. Lupus nephritis yang diinduksi pristane di Nrf2? /? tikus yang diobati dengan sulforaphane memiliki kerusakan ginjal yang parah dan perubahan patologis serta peningkatan ekspresi iNOS dan aktivasi NF-? B dibandingkan dengan WT, menunjukkan bahwa Nrf2 memperbaiki lupus nephritis dengan menghambat jalur pensinyalan NF-? B dan membersihkan ROS [94 ]. Aktivitas NF-? B juga terjadi ketika sel diperlakukan dengan penginduksi Nrf2 bersama-sama dengan LPS dan TNF- ?. Sebagai contoh, turunan chalcone sintetis menghambat aktivasi NF-? B yang diinduksi TNF -? - baik secara langsung maupun tidak langsung dan sebagian melalui induksi ekspresi HO-1 dalam sel HT-29 epitel usus manusia [62]. Supresi translokasi NF-? B dan aktivitas pengikatan DNA serta penekanan ekspresi iNOS dalam hepatosit ditemukan ketika tikus F344 diobati dengan 3H-1,2-dithiole-3-thione (D3T) [95]. Setelah pengobatan bersama dengan sulforaphane dan LPS, ekspresi iNOS, COX-2, dan TNF-? dalam 264.7 makrofag mentah diatur ke bawah, menunjukkan bahwa sulforaphane memiliki aktivitas anti-inflamasi melalui penghambatan pengikatan DNA NF-? B [96]. Meskipun beberapa studi eksperimental telah dilakukan untuk menjelaskan hubungan antara jalur Nrf2 dan NF-? B, hasil yang bertentangan tetap ada. Kedua peraturan positif dan negatif telah dilaporkan antara Nrf2 dan NF-kB [97]. Biasanya, elektrofisi kemopreventif 3H-1,2-dithiole-3-thione, sulforaphane dan Triterpenoid CDDO-Me mengaktifkan Nrf2 dengan menghambat NF-kB dan gen yang diregulasi [98], [99], [100]. Sebaliknya, beberapa agen atau kondisi seperti ROS, LPS, aliran tegangan geser, LDL teroksidasi, dan asap rokok telah terbukti meningkatkan aktivitas Nrf2 dan NF-kB [97]. Selain itu, penelitian in vivo telah mengungkapkan bahwa aktivitas NF-kB menurun pada hati yang diisolasi dari Nrf2? /? tikus dan aktivitas pengikatan NF-? B lebih rendah di Nrf2? /? daripada di Nrf2 + / + mice [101]. Namun, sel endotel aorta manusia yang diobati dengan vektor adenoviral Nrf2 menghambat gen hilir NF-? B tanpa mempengaruhi aktivitas NF-? B [8].
Gambar 4 Loop regulasi Nrf2 dan NF-? B. Jalur Nrf2 menghambat aktivasi NF-? B dengan mencegah degradasi I? B-? dan meningkatkan ekspresi HO-1 dan pertahanan antioksidan yang menetralkan ROS dan bahan kimia detoksifikasi. Akibatnya, aktivasi NF-? B terkait ROS ditekan. Demikian juga, transkripsi yang dimediasi NF-? B mengurangi aktivasi Nrf2 dengan mengurangi ADALAHTranskripsi gen dan protein pengikat CREB bebas dengan bersaing dengan Nrf2 untuk CBP. Selain itu, NF-? B meningkatkan perekrutan histone deacetylase (HDAC3) ke wilayah ARE dan karenanya aktivasi transkripsi Nrf2 dicegah.
Aktivasi jalur pensinyalan Nrf2 memainkan peran utama dalam ekspresi enzim dan gen yang terlibat dalam detoksifikasi oksidan reaktif dengan meningkatkan kapasitas antioksidan sel dalam tubuh manusia. Sementara banyak studi penelitian tersedia saat ini, mekanisme pengaturan dalam aktivasi Nrf2 tidak sepenuhnya dipahami. Kemungkinan peran jalur pensinyalan Nrf2 dalam pengobatan peradangan juga telah ditemukan. Dr Alex Jimenez DC, CCST Insight
Peran Nrf2 pada Penyakit Inflamasi
Studi in vivo telah menunjukkan bahwa Nrf2 memainkan peran penting dalam penyakit radang yang mempengaruhi sistem yang berbeda; ini termasuk gastritis, radang usus besar, radang sendi, pneumonia, kerusakan hati, penyakit kardiovaskular, penyakit neurodegeneratif, dan kerusakan otak. Dalam studi ini, Nrf2? /? hewan menunjukkan gejala peradangan dan kerusakan jaringan yang lebih parah daripada hewan WT. Oleh karena itu, diyakini bahwa jalur sinyal Nrf2 memiliki efek perlindungan pada penyakit inflamasi. Instalasi intra-trakea dari porcine pankreas elastase menginduksi penyakit paru obstruktif kronik, terutama emfisema. Tikus defisien-Nrf2 sangat rentan terhadap emfisema, dan penurunan ekspresi HO-1, PrxI, dan gen antiprotease SLPI terjadi pada makrofag alveolar. Nrf2 dianggap sebagai pengatur utama dalam sistem pertahanan makrofag yang dimediasi terhadap cedera paru [102]. Tikus dengan defisiensi Nrf2 dengan emfisema yang diinduksi oleh paparan asap tembakau untuk bulan 6 menunjukkan peningkatan peradangan bronchoalveolar, peningkatan ekspresi penanda stres oksidatif di alveoli, dan peningkatan apoptosis septum sel alveolar, menunjukkan bahwa Nrf2 bekerja melawan emfisema yang diinduksi tembakau melalui peningkatan ekspresi antioksidan. gen [102], [103]. Dengan gangguan Nrf2, peradangan saluran napas alergen-mediated dan asma menggunakan ovalbumin kompleks menunjukkan peningkatan inflamasi saluran napas, reaktivitas hiper-jalan, hiperplasia sel goblet, dan tingkat tinggi Th2 di bronchoalveolar lavage dan splenocytes, sedangkan jalur signaling Nrf2-dimediasi membatasi eosinofilia jalan napas , hipersekresi lendir, dan hiper-reaktivitas saluran napas serta menginduksi banyak gen antioksidan yang mencegah perkembangan asma [104]. Injeksi karagenan ke dalam rongga pleura menginduksi pleuritis, dan akumulasi 15d-PGJ2 pada sel inflamasi Nrf2 terbatas pada makrofag peritoneum tikus. Selama fase awal peradangan, 15d-PGJ2 mengaktifkan Nrf2 dan mengatur proses inflamasi melalui induksi HO-1 dan PrxI. Sebuah penelitian juga menunjukkan bahwa COX-2 memiliki efek anti-inflamasi pada fase awal dengan produksi 15d-PGJ2 [105]. Pemberian oral 1% dekstran sulfat natrium untuk minggu 1 menginduksi kolitis yang terkait dengan perubahan histologis yang termasuk pemendekan crypts dan infiltrasi sel-sel inflamasi pada jaringan usus besar. Untuk melindungi integritas usus dalam kolitis, Nrf2 dapat memainkan peran penting dengan mengatur sitokin pro-inflamasi dan menginduksi enzim detoksifikasi fase II [51]. Dalam model tikus Nrf2-knockout dari sepsis paru yang diinduksi LPS, aktivitas NF-? B mengatur pengaruh sitokin inflamasi seperti COX-2, IL-113, IL-6, dan TNF? yang penting untuk memulai dan mempromosikan peradangan [60]. Nrf2 mengurangi kerusakan inflamasi dengan mengatur faktor-faktor inflamasi ini. Dalam model-model peradangan akut ini, peningkatan regulasi enzim antioksidan, sitokin pro-inflamasi, dan mediator oleh jalur pensinyalan Nrf2 mengurangi cedera inflamasi pada hewan WT. Menariknya, ini juga telah dilaporkan pada tikus Nrf2-knockout di mana gejala yang nyata diperberat dibandingkan dengan tikus WT.
Penelitian Nrf2-Dependent Anti-Inflammatory Drugs
Singkatnya, kami telah mendiskusikan eksperimen yang menunjukkan bahwa jalur sinyal Nrf2 memainkan peran pengatur di banyak area peradangan, sehingga agen anti-inflamasi Nrf2 bergantung penting untuk pengobatan penyakit inflamasi.
Tanaman telah menjadi sumber yang sangat kaya senyawa yang mengaktifkan faktor transkripsi Nrf2, yang mengarah ke up-regulasi gen cytoprotective. Baru-baru ini, beberapa penelitian dilakukan untuk menyelidiki efek dari berbagai agen anti-inflamasi, sebagian besar berasal dari tumbuhan. Misalnya, kurkumin adalah bahan aktif kunyit dan juga ditemukan dalam jumlah kecil pada jahe; isothiocyanate, khususnya phenylisothiocyanate berasal dari brokoli, seledri, dan sayuran lainnya; dan anthocyanin berasal dari buah beri dan anggur [124]. Penelitian telah menunjukkan bahwa semua agen ini tidak hanya antioksidan yang baik tetapi juga memiliki efek anti-inflamasi yang kuat melalui induksi Nrf2 [125], [126]. Oleh karena itu, pengembangan aktivator Nrf2 anti-inflamasi baru dari ekstrak tumbuhan telah menarik banyak minat dalam penelitian medis.
Dalam beberapa tahun terakhir, banyak percobaan pada hewan telah dilakukan untuk mengkonfirmasi aksi senyawa ini. Artesunat digunakan terutama untuk malaria berat, malaria otak, dan penyakit autoimun rematik; itu juga efektif pada cedera paru septik. Artesunat mengaktifkan ekspresi Nrf2 dan HO-1, dan yang terakhir mengurangi aliran sitokin dan leukosit pro-inflamasi ke dalam jaringan untuk mencegah peradangan [127]. Isovitexin, diekstrak dari kulit beras Oryza sativa, dianggap memiliki sifat anti-inflamasi dan antioksidan; ia memainkan peran protektif terhadap cedera paru akut yang diinduksi LPS dengan mengaktifkan jalur Nrf2 / HO-1 dan menghambat MAPK dan NF-? B [128]. Fimasartan, penghambat reseptor angiotensin II yang baru populer yang bekerja pada sistem renin-angiotensin, mengurangi tekanan darah; menggunakan fimasartan untuk mengobati tikus dengan obstruksi ureter unilateral yang diinduksi melalui pembedahan mengurangi stres oksidatif, peradangan, dan fibrosis melalui pengaturan Nrf2 dan jalur antioksidan serta menghambat RAS dan MAPKs [129]. Sappanone didistribusikan secara luas di Asia Tenggara, di mana ia digunakan sebagai obat anti influenza, anti alergi, dan pelindung saraf; itu mengaktifkan Nrf2 dan menghambat NF-? B dan mungkin bermanfaat dalam pengobatan penyakit terkait Nrf2- dan / atau NF-? B [130]. Bixin yang diekstrak dari biji Bixin orellana digunakan untuk penyakit infeksi dan inflamasi di Meksiko dan Amerika Selatan; itu mengurangi mediator inflamasi, kebocoran kapiler alveolar, dan kerusakan oksidatif dengan cara yang bergantung pada Nrf2 untuk mengurangi cedera paru yang diinduksi ventilasi dan mengembalikan morfologi paru yang normal [131]. Senyawa tanaman lain, seperti epigallocatechin gallate, sulforaphane, resveratrol, likopen, dan ekstrak teh hijau memiliki efek terapeutik pada penyakit inflamasi melalui jalur pensinyalan Nrf2 [132], [133], [134]. Baru-baru ini fitokimia lain, eriodictyol, yang ada dalam buah jeruk, telah dilaporkan memiliki efek antiinflamasi dan antioksidan pada cedera ginjal yang diinduksi cisplatin dan cedera paru akut yang diinduksi sepsis dengan mengatur Nrf2, menghambat NF-? B, dan ekspresi sitokin di makrofag [135], [136]. Namun, banyak fitokimia yang menjanjikan untuk pencegahan dan pengobatan berbagai penyakit manusia, dan beberapa telah memasuki tahap uji klinis (Tabel 2).
Senyawa tanaman ini mengaktifkan jalur pensinyalan Nrf2 terutama dalam bentuk bahan elektrofilik yang memodifikasi residu sistein Keap1, yang mengarah ke ikatan Nrf2 nuklir bebas dengan ARE, yang menghasilkan aktivasi transkripsi gen yang sesuai.
Sulforaphane dan Dampaknya pada Kanker, Kematian, Penuaan, Otak dan Perilaku, Penyakit Jantung & Lainnya
Isothiocyanate adalah beberapa senyawa tanaman yang paling penting yang bisa Anda dapatkan dalam diet Anda. Dalam video ini saya membuat kasus yang paling komprehensif untuk mereka yang pernah dibuat. Rentang perhatian yang pendek? Lewati ke topik favorit Anda dengan mengklik salah satu poin waktu di bawah ini. Garis waktu penuh di bawah ini.
Bagian utama:
00: 01: 14 - Kanker dan kematian
00: 19: 04 - Penuaan
00: 26: 30 - Otak dan perilaku
00: 38: 06 - Rekap terakhir
00: 40: 27 - Dosis
Lini waktu penuh:
00: 00: 34 - Pengantar sulforaphane, fokus utama dari video.
00: 01: 14 - konsumsi sayuran Cruciferous dan pengurangan dalam semua penyebab kematian.
00: 02: 12 - Risiko kanker prostat.
00: 02: 23 - Risiko kanker kandung kemih.
00: 02: 34 - Kanker paru-paru berisiko pada perokok.
00: 02: 48 - Risiko kanker payudara.
00: 03: 13 - Hipotetis: bagaimana jika Anda sudah menderita kanker? (intervensi)
00: 03: 35 - Mekanisme yang masuk akal mengemudi kanker dan data asosiatif kematian.
00: 04: 38 - Sulforaphane dan kanker.
00: 05: 32 - Hewan bukti yang menunjukkan efek kuat dari ekstrak kecambah brokoli pada perkembangan tumor kandung kemih pada tikus.
00: 06: 06 - Pengaruh suplementasi langsung sulforaphane pada pasien kanker prostat.
00: 07: 09 - Bioakumulasi metabolit isothiocyanate dalam jaringan payudara yang sebenarnya.
00: 08: 32 - Penghambatan sel induk kanker payudara.
00: 08: 53 - Pelajaran sejarah: brassica ditetapkan memiliki sifat-sifat kesehatan bahkan di Roma kuno.
00: 09: 16 - Kemampuan Sulforaphane untuk meningkatkan ekskresi karsinogen (benzena, akrolein).
00: 09: 51 - NRF2 sebagai saklar genetik melalui elemen respons antioksidan.
00: 10: 10 - Bagaimana aktivasi NRF2 meningkatkan ekskresi karsinogen melalui glutathione-S-conjugates.
00: 10: 34 - kubis Brussel meningkatkan glutathione-S-transferase dan mengurangi kerusakan DNA.
00: 11: 20 - Broccoli sprout drink meningkatkan ekskresi benzena oleh 61%.
00: 13: 31 - Broccoli sprout homogenate meningkatkan enzim antioksidan di saluran napas bagian atas.
00: 15: 45 - konsumsi sayuran Cruciferous dan kematian penyakit jantung.
00: 16: 55 - Bubuk tunas brokoli meningkatkan lipid darah dan risiko penyakit jantung secara keseluruhan pada penderita diabetes tipe 2.
00: 19: 04 - Awal dari bagian penuaan.
00: 19: 21 - Diet yang diperkaya Sulforaphane meningkatkan masa hidup kumbang dari 15 ke 30% (dalam kondisi tertentu).
00: 20: 34 - Pentingnya peradangan rendah untuk umur panjang.
00: 22: 05 - Sayuran dan tunas kecambah brokoli tampaknya mengurangi beragam penanda inflamasi pada manusia.
00: 23: 40 - Rekap video pertengahan: kanker, bagian penuaan
00: 24: 14 - Studi pada tikus menunjukkan sulforaphane dapat meningkatkan fungsi imun adaptif di usia tua.
00: 25: 18 - Sulforaphane meningkatkan pertumbuhan rambut pada model tikus botak. Gambar di 00: 26: 10.
00: 26: 30 - Awal dari bagian otak dan perilaku.
00: 27: 18 - Pengaruh ekstrak kecambah brokoli pada autisme.
00: 27: 48 - Pengaruh glucoraphanin pada skizofrenia.
00: 28: 17 - Mulai dari diskusi depresi (mekanisme dan studi yang masuk akal).
00: 31: 21 - Studi mouse menggunakan 10 model yang berbeda dari depresi yang diinduksi stres menunjukkan sulforaphane sama efektifnya dengan fluoxetine (prozac).
00: 32: 00 - Studi menunjukkan konsumsi langsung glukoraphanin pada tikus juga efektif mencegah depresi dari model stres kekalahan sosial.
00: 33: 01 - Awal dari bagian neurodegenerasi.
00: 33: 30 - Sulforaphane dan penyakit Alzheimer.
00: 33: 44 - Sulforaphane dan penyakit Parkinson.
00: 33: 51 - Sulforaphane dan penyakit Hungtington.
00: 34: 13 - Sulforaphane meningkatkan protein heat shock.
00: 34: 43 - Awal dari bagian cedera otak traumatis.
00: 35: 01 - Sulforaphane disuntikkan segera setelah TBI meningkatkan daya ingat (studi pada tikus).
00: 35: 55 - Sulforaphane dan plastisitas neuronal.
00: 36: 32 - Sulforaphane meningkatkan pembelajaran dalam model diabetes tipe II pada tikus.
00: 37: 19 - Sulforaphane dan duchenne muscular dystrophy.
00: 37: 44 - Myostatin inhibition dalam sel-sel satelit otot (in vitro).
00: 38: 06 - Rekap video-ulang: mortalitas dan kanker, kerusakan DNA, stres oksidatif dan peradangan, ekskresi benzena, penyakit kardiovaskular, diabetes tipe II, efek pada otak (depresi, autisme, skizofrenia, neurodegenerasi), jalur NRF2.
00: 40: 27 - Pikiran tentang mencari tahu kecambah brokoli atau sulforaphane.
00: 41: 01 - Anekdot saat bertumbuh di rumah.
00: 43: 14 - Pada suhu memasak dan aktivitas sulforaphane.
00: 43: 45 - Konversi bakteri usus dari sulforaphane dari glucoraphanin.
00: 44: 24 - Suplemen bekerja lebih baik ketika dikombinasikan dengan myrosinase aktif dari sayuran.
00: 44: 56 - Teknik memasak dan sayuran silangan.
00: 46: 06 - Isothiocyanate sebagai goitrogens.
Kesimpulan
Saat ini, banyak penelitian telah difokuskan pada peran jalur pensinyalan Nrf2 / Keap1 / ARE dalam peradangan. Di antara enzim yang diregulasi oleh Nrf2, HO-1 adalah salah satu enzim respons stres yang representatif. HO-1 memiliki sifat anti-inflamasi dan antioksidan yang menonjol. Secara umum, jalur pensinyalan Nrf2 juga secara negatif mengatur sitokin, faktor pelepas kemokin, MMPs, dan mediator inflamasi lainnya COX-2 dan produksi iNOS, yang secara langsung atau tidak langsung mempengaruhi jalur NF-kB dan MAPK yang relevan dan jaringan lain yang mengontrol peradangan. Disarankan bahwa jalur pensinyalan Nrf2 dan NF-? B berinteraksi untuk mengatur transkripsi atau fungsi protein target hilir. Penekanan atau inaktivasi aktivitas transkripsi yang dimediasi NF-? B melalui Nrf2 kemungkinan besar terjadi pada fase awal inflamasi, karena NF-? B mengatur sintesis de novo dari serangkaian mediator pro-inflamasi. Namun demikian, masih terdapat beberapa keterbatasan dalam penelitian seperti apakah terdapat hubungan antara Nrf2 dengan jalur pensinyalan lainnya seperti JAK / STAT, pentingnya aktivator Nrf2 saat ini yang berasal dari sumber tumbuhan alami pada inflamasi, dan bagaimana meningkatkan aktivitas biologis. dan meningkatkan penargetan senyawa ini. Ini membutuhkan validasi eksperimental lebih lanjut.
Selain itu, jalur pensinyalan Nrf2 dapat mengatur > 600 gen [163], di mana > 200 mengkode protein sitoprotektif [164] yang juga terkait dengan peradangan, kanker, penyakit neurodegeneratif, dan penyakit utama lainnya [165]. Semakin banyak bukti yang menunjukkan bahwa, jalur pensinyalan Nrf2 dideregulasi di banyak kanker, menghasilkan ekspresi yang menyimpang dari baterai gen yang bergantung pada Nrf2. Selain itu, peradangan memainkan peran utama dalam penyakit terkait stres oksidatif terutama pada kanker. Penerapan beberapa aktivator Nrf2 untuk melawan peradangan dapat mengakibatkan ekspresi menyimpang dari gen hilir Nrf2 yang menginduksi onkogenesis dan resistensi terhadap kemoterapi dan/atau terapi radio. Oleh karena itu, aktivator Nrf2 yang sangat spesifik dapat dikembangkan untuk meminimalkan efek pleiotropiknya. Beberapa aktivator Nrf2 telah menunjukkan peningkatan yang signifikan dari fungsi anti-inflamasi pada penyakit terkait stres oksidatif. Contoh aktivator Nrf2 terbaik yang disetujui oleh FDA dan banyak digunakan untuk pengobatan penyakit inflamasi seperti Multiple sclerosis (MS) adalah dimetil fumarat. Tecfidera� (nama terdaftar dimetil fumarat oleh Biogen) digunakan secara efektif untuk mengobati bentuk sklerosis multipel yang kambuh pada sejumlah besar pasien [152]. Namun, kemanjuran menggunakan aktivator Nrf2 untuk mengobati penyakit inflamasi memerlukan validasi lebih lanjut untuk menghindari efek merusak dari Nrf2. Oleh karena itu, pengembangan terapi untuk aktivitas anti-inflamasi yang dimediasi oleh Nrf2 dapat memiliki dampak klinis yang signifikan. Studi yang sedang berlangsung dari jalur pensinyalan Nrf2 di seluruh dunia dikhususkan untuk mengembangkan agen terapeutik yang sangat bertarget untuk mengendalikan gejala peradangan, dan untuk mencegah dan mengobati kanker serta neurodegeneratif dan penyakit utama lainnya.
Kesimpulannya, Nrf2 merasakan tingkat stres oksidatif dalam tubuh manusia dan akhirnya membantu mempromosikan pengaturan enzim dan gen antioksidan dan detoksifikasi. Karena peradangan kronis yang disebabkan oleh peningkatan tingkat stres oksidatif telah dikaitkan dengan penyakit neurodegeneratif, Nrf2 dapat memainkan peran penting dalam pengobatan masalah kesehatan seperti penyakit Alzheimer, antara lain. Cakupan informasi kami terbatas pada masalah chiropraktik dan kesehatan tulang belakang. Untuk membahas pokok bahasan ini, silakan bertanya kepada Dr. Jimenez atau hubungi kami di 915-850-0900 .
Diskusi Topik Tambahan: Menghilangkan Nyeri Lutut tanpa Pembedahan
Nyeri lutut adalah gejala umum yang dapat terjadi karena berbagai cedera dan / atau kondisi lutut, termasuk cedera olahraga. Lutut adalah salah satu sendi yang paling kompleks dalam tubuh manusia karena terdiri dari persimpangan empat tulang, empat ligamen, berbagai tendon, dua menisci, dan tulang rawan. Menurut American Academy of Family Physicians, penyebab paling umum dari nyeri lutut termasuk subluksasi patela, tendinitis patela atau lutut pelompat, dan penyakit Osgood-Schlatter. Meskipun nyeri lutut paling mungkin terjadi pada orang berusia di atas 60 tahun, nyeri lutut juga dapat terjadi pada anak-anak dan remaja. Nyeri lutut dapat diobati di rumah dengan mengikuti metode RICE, namun cedera lutut yang parah mungkin memerlukan perhatian medis segera, termasuk perawatan chiropraktik. �
Penyakit neurodegeneratif, seperti penyakit Alzheimer dan penyakit Parkinson, memengaruhi jutaan orang di seluruh dunia. Berbagai pilihan pengobatan tersedia untuk mengobati gejala beberapa penyakit neurodegeneratif walaupun hasilnya sering terbatas. Studi penelitian telah menemukan bahwa stres oksidatif yang disebabkan oleh faktor internal dan eksternal dapat menjadi penyebab perkembangan penyakit neurodegeneratif. Itu faktor transkripsi, Nrf2, telah bertekad untuk berfungsi sebagai mekanisme pertahanan utama terhadap stres oksidatif. Tujuan artikel di bawah ini adalah untuk menunjukkan efek Nrf2 pada penyakit neurodegeneratif.
Modulasi dari Proteostasis oleh Faktor Transkripsi NRF2
Penyakit neurodegeneratif terkait dengan akumulasi agregat protein spesifik, menunjukkan hubungan intim antara otak yang terluka dan hilangnya proteostasis. Proteostasis mengacu pada semua proses di mana sel mengontrol kelimpahan dan melipat dari proteome berkat jaringan luas yang mengintegrasikan pengaturan jalur sinyal, ekspresi gen dan sistem degradasi protein. Ulasan ini mencoba merangkum temuan yang paling relevan tentang modulasi transkripsi dari proteostasis yang diberikan oleh faktor transkripsi NRF2 (faktor nuklir (2 yang diturunkan dari erythroid)) seperti 2). NRF2 telah secara klasik dianggap sebagai pengatur utama dari respon sel antioksidan, meskipun saat ini muncul sebagai komponen kunci dari mesin transduksi untuk mempertahankan proteostasis. Seperti yang akan kita diskusikan, NRF2 dapat dibayangkan sebagai hub yang mengkompilasi sinyal-sinyal darurat yang berasal dari akumulasi protein yang gagal melipat untuk membangun respon transkripsional yang terkoordinasi dan dapat dibangkitkan. Hal ini dicapai oleh fungsi NRF2 terkait dengan kontrol gen yang terlibat dalam pemeliharaan fisiologi retikulum endoplasma, proteasome dan autophagy.
Kata kunci:Penyakit neurodegeneratif, Respon protein yang tidak dilipat, Proteasome, Ubiquitin, Autophagy, Stres oksidatif
Nuclear Factor (erythroid-derived 2) -seperti 2 (NRF2) adalah protein dasar-leusin-ritsleting yang dianggap saat ini sebagai pengatur utama homeostasis seluler. Ini mengontrol ekspresi basal dan ekspresi yang diinduksi-stres dari gen 250 yang berbagi secara umum peningkat cis-acting yang disebut elemen respon antioksidan (ADALAH) [1], [2], [3], [4], [5]. Gen-gen ini berpartisipasi dalam fase I, II dan III reaksi detoksifikasi, glutathione dan peroxiredoxin / metabolisme thioredoxin, produksi NADPH melalui jalur pentosa fosfat dan enzim malat, oksidasi asam lemak, metabolisme besi, dan proteostasis [6]. Dengan adanya fungsi-fungsi sitoprotektif yang luas ini, ada kemungkinan bahwa satu pukulan farmakologi dalam NRF2 dapat mengurangi efek dari penyebab utama penyakit kronis, termasuk stres oksidatif, inflamasi dan proteotoksik. Peran NRF2 dalam modulasi pertahanan antioksidan dan resolusi peradangan telah dibahas dalam berbagai penelitian (ditinjau dalam [7]). Di sini, kami akan fokus pada perannya dalam proteostasis, yaitu kontrol homeostatik sintesis protein, lipat, perdagangan dan degradasi. Contoh akan diberikan dalam konteks penyakit neurodegeneratif.
Hilangnya Pengaruh Proteostasis NRF2 Kegiatan di Neurodegenerative Diseases
Ciri umum penyakit neurodegeneratif adalah terjadinya agregasi yang menyimpang dari beberapa protein. Dengan demikian, agregat protein yang salah lipatan dari? -Synuclein (? -SYN) ditemukan pada penyakit Parkinson (PD), plak? -Amyloid (A?) Dan kusut neurofibrillary TAU hiperfosforilasi pada penyakit Alzheimer (AD), huntingtin (Htt) di Penyakit Huntington (HD), superoksida dismutase 1 (SOD1) dan protein pengikat DNA TAR 43 (TDP-43) pada amyotrophic lateral sclerosis (ALS), protein prion (PrP) pada ensefalopati spongiform, dll. Agregat protein dapat berdampak pada beberapa penyakit. jalur seluler, yang pada gilirannya dapat memengaruhi tingkat dan aktivitas NRF2.
Lapisan Peraturan yang Berbeda Mengontrol Aktivitas NRF2 dengan Ketat
Di bawah kondisi fisiologis, sel menunjukkan tingkat protein NRF2 yang rendah karena perputarannya yang cepat. Menanggapi rangsangan yang berbeda, protein NRF2 terakumulasi, memasuki nukleus dan meningkatkan transkripsi gen yang mengandung ARE. Oleh karena itu, manajemen tingkat protein NRF2 adalah titik kunci yang harus mengintegrasikan sinyal input positif dan negatif. Seperti yang akan kita bahas lebih lanjut, NRF2 diaktifkan oleh mekanisme tumpang tindih yang beragam untuk mengatur respons yang cepat dan efisien tetapi di sisi lain NRF2 dapat dihambat, mungkin dalam fase kedua, untuk mematikan responsnya.
Dari sudut pandang klasik, aktivasi NRF2 telah dianggap sebagai konsekuensi dari respon seluler terhadap senyawa oksidan atau elektrofilik. Dalam hal ini, ubiquitin E3 ligase adapter Kelch-like ECH-related protein 1 (KEAP1) memainkan peran penting. Rincian molekuler akan dibahas lebih lanjut di Bagian 4.1. Singkatnya, KEAP1 bertindak sebagai sensor redoks karena residu sistein kritis yang menyebabkan NRF2 di mana-mana dan degradasi proteasomal. Selain modulasi klasik ini, NRF2 sangat diatur oleh peristiwa pensinyalan. Memang, kinase yang berbeda telah terbukti memfosforilasi dan mengatur NRF2. Misalnya, NRF2 dapat difosforilasi oleh mitogen aktivasi protein kinase (MAPKs), meskipun kontribusinya terhadap aktivitas NRF2 masih belum jelas [8], [9], [10], [11]. PKA kinase serta beberapa isozim PKC [12], CK2 [13] atau Fyn [14] fosforilat NRF2 memodifikasi stabilitasnya. Pekerjaan sebelumnya dari kelompok kami melaporkan bahwa glikogen sintase kinse-3? (GSK-3?) Menghambat NRF2 dengan eksklusi nuklir dan degradasi proteasomal [15], [25], [26], [27], [28], [29], [30]. Rincian molekuler akan dibahas di Bagian 4.1. Selanjutnya, NRF2 diserahkan kepada jenis regulasi lain. Misalnya, asetilasi NRF2 oleh CBP / p300 meningkatkan aktivitasnya [17], sementara itu dihambat oleh miR153, miR27a, miR142-5p, dan miR144 [16], atau dengan metilasi pulau sitosin-guanin (CG) dalam promotor NRF2 [18].
Dampak Agregat Protein pada Mekanisme Regulasi NRF2
Pada bagian ini kita akan fokus pada bagaimana akumulasi dari protein yang gagal melipat dapat mempengaruhi aktivitas NRF2 yang menyediakan beberapa jalur yang disebutkan di atas sebagai contoh ilustratif. Pertama, kita perlu mempertimbangkan bahwa akumulasi protein telah dikaitkan erat dengan kerusakan oksidatif. Memang, akumulasi protein dan agregasi yang gagal melipatgandakan menginduksi produksi abnormal spesies oksigen reaktif (ROS) dari mitokondria dan sumber lain [19]. Sebagaimana disebutkan di atas, ROS akan memodifikasi cysteines redox-sensitif KEAP1 yang mengarah pada pelepasan, stabilisasi dan lokalisasi nuklir NRF2.
Mengenai proteinopati, contoh kejadian pensinyalan yang tidak diatur yang dapat mempengaruhi NRF2 disediakan oleh hiperaktivasi GSK-3? dalam iklan. GSK-3 ?, juga dikenal sebagai TAU kinase, berpartisipasi dalam fosforilasi protein terkait mikrotubulus ini, menghasilkan agregasi, pembentukan kusut neurofibrillary dan gangguan transpor aksonal (ditinjau dalam [20]). Di sisi lain, GSK-3? secara dramatis mengurangi tingkat dan aktivitas NRF2 seperti yang disebutkan di atas. Meskipun tidak diterima secara luas, kaskade amiloid menyatakan bahwa racun A? oligomer meningkatkan GSK-3? aktivitas bersama dengan hiperfosforilasi TAU dan kematian neuron [21], [22]. Ada berbagai model untuk menjelaskan bagaimana A? menyukai GSK3-? aktivitas. Misalnya, A? mengikat reseptor insulin dan menghambat jalur pensinyalan PI3K dan AKT, yang penting untuk mempertahankan GSK-3? dinonaktifkan oleh fosforilasi pada residu N-terminal Ser9 [23]. Di sisi lain, ekstraseluler A? berinteraksi dengan reseptor Frizzled, memblokir pensinyalan WNT [24] dan sekali lagi menghasilkan pelepasan GSK-3 yang aktif. Singkatnya, A? akumulasi menyebabkan hiperaktivasi abnormal GSK-3?, sehingga mengganggu respons NRF2 yang sesuai.
Sebagaimana dibahas dalam bagian berikut, protein yang gagal melipat menyebabkan aktivasi PERK dan MAPKs, yang pada gilirannya mengatur NRF2 [31], [8], [9], [10], [11]. Selain itu, aktivitas CBP / p300 yang tidak diregulasi telah dilaporkan pada beberapa proteinopathies [32] dan penurunan global dalam metilasi DNA pada otak AD juga ditunjukkan [33], oleh karena itu memberikan alasan untuk mengeksplorasi relevansi dari temuan ini dalam regulasi NRF2.
Kami dan orang lain telah mengamati pada nekropsi pasien PD dan AD peningkatan kadar protein NRF2 dan beberapa targetnya, seperti heme oxygenase 1 (HMOX1), NADPH quinone oxidase 1 (NQO1), p62, dll., Baik oleh immunoblot dan oleh imunohistokimia [34], [35], [36], [37], [38], [39]. Pengaturan NRF2 pada penyakit ini ditafsirkan sebagai upaya gagal otak yang sakit untuk memulihkan nilai homeostatik. Namun, penelitian lain menunjukkan bahwa NRF2 sebagian besar terlokalisasi di sitoplasma neuron hippokampus AD, menunjukkan aktivitas transkripsi NRF2 berkurang di otak [40]. Bisa dibayangkan bahwa perbedaan pengamatan ini terkait dengan perubahan faktor-faktor yang mengontrol NRF2 sepanjang tahapan progresif neurodegenerasi.
Tiga sistem utama berkontribusi terhadap proteostasis, yaitu respon protein yang terbentang (UPR), sistem proteasome ubiquitin (UPS) dan autophagy. Selanjutnya, kami menyajikan bukti untuk membayangkan NRF2 sebagai hub yang menghubungkan sinyal darurat yang diaktifkan oleh agregat protein dengan mesin turunan protein.
NRF2 Berpartisipasi dalam Respons Protein yang Tidak Terungkap (UPR)
Aktivasi NRF2 dalam Tanggapan ke UPR
Pelipatan protein oksidatif di UGD didorong oleh sejumlah jalur berbeda, yang paling terkonservasi di antaranya melibatkan protein disulfida-isomerase (PDI) dan oksidoreduktin endoplasma sulfhidril oksidase 1 (ERO1? Dan ERO1? Pada mamalia) sebagai donor disulfida. Secara singkat, PDI mengkatalisis pembentukan dan kerusakan ikatan disulfida antara residu sistein dalam protein, saat mereka melipat, karena reduksi dan oksidasi asam amino sisteinnya sendiri. PDI didaur ulang oleh aksi dari enzim rumah tangga ERO1, yang memperkenalkan kembali ikatan disulfida ke dalam PDI [41]. Oksigen molekuler adalah akseptor elektron terminal ERO1, yang menghasilkan jumlah stoikiometri hidrogen peroksida untuk setiap ikatan disulfida yang dihasilkan [42]. Peroksidase (PRX4) dan glutathione peroksidase (GPX7 dan GPX8) adalah enzim kunci untuk mengurangi hidrogen peroksida di ER. Ketika sistem oksido-reduktif ini tidak bekerja dengan baik, akumulasi abnormal dari protein yang salah lipatan terjadi di UGD dan satu set sinyal bernama respon protein yang tidak dilipat (UPR) ditransmisikan ke sitoplasma dan nukleus untuk membangun kembali homeostasis ER [43]. Tiga protein yang berhubungan dengan membran telah diidentifikasi untuk merasakan tekanan ER pada eukariota: mengaktifkan faktor transkripsi 6 (ATF6), ER eIF2 pankreas? kinase (PERK, juga beruntai ganda RNA-teraktivasi protein kinase-seperti ER kinase), dan inositol-membutuhkan kinase1 (IRE1). Domain luminal dari setiap sensor terikat pada pendamping 78 kDa yang disebut protein yang diatur glukosa (GRP78 / BIP). BIP berdisosiasi pada tekanan ER untuk mengikat protein yang tidak terlipat, yang mengarah ke aktivasi tiga sensor [44].
NRF2 dan homologinya NRF1, juga terkait dengan respon antioksidan, berpartisipasi dalam transduksi UPR ke nukleus. Dalam kasus NRF1, protein ini terletak di membran ER dan mengalami translokasi nuklir pada deglikosilasi atau pembelahan. Kemudian, aktivasi UPR mengarah pada pemrosesan NRF1 dan akumulasi nuklir dari fragmen yang dihasilkan dalam kompartemen nuklir. Namun, kemampuan untuk mentransmisikan gen yang mengandung ARE dari fragmen NRF1 ini masih dalam pembahasan [45].
Glover-Cutter dan rekan kerja menunjukkan aktivasi ortholog NRFPNUMX dari C. elegans, SKN-2, dengan berbagai ER stressor. Peningkatan ekspresi SKN-1 tergantung pada mediator UPR yang berbeda, termasuk IRE1 atau PERK cacing orthologues [1]. Dalam sel PERK-defisiensi, sintesis protein terganggu menyebabkan akumulasi peroksida endogen dan apoptosis [46] berikutnya. Efektor yang digunakan oleh PERK untuk melindungi ER dari peroksida ini mungkin NRF47, karena telah dilaporkan bahwa PERK memfosforilasi NRF2 di Ser2, sehingga mencegah degradasi oleh KEAP40 [1]. Induksi ASK31 juga cenderung berperan dalam rute ini melalui aksi kinase yang dimediasi TRAF1 dari IRE2 [1]. Meskipun peran MAPK dalam regulasi NRF48 masih kontroversial, baru-baru ini disarankan bahwa jalur IRE2-TRAF1-ASK2-JNK mungkin mengaktifkan NRF1 [2] (Gambar 49). Menariknya, pada C. elegans dan sel manusia, bukti baru menunjukkan bahwa cysteine sulfenylation dari IRE1 kinase pada loop aktivasi menghambat IRE1-mediated UPR dan memulai respon antioksidan p1 yang digerakkan oleh NRF38. Data menunjukkan bahwa IRE2 memiliki fungsi kuno sebagai sentinel sitoplasma yang mengaktifkan p1 dan NRF38 [2].
Gambar 1 Peraturan NRF2 oleh UPR. Akumulasi protein yang tidak terbuka atau salah dalam retikulum endoplasma dapat memicu respon protein yang tidak terbuka (UPR). Pertama, BIP pendamping dilepaskan dari domain intraluminal dari sensor ER IRE1 dan PERK untuk mengikat protein yang tidak dilipat / gagal melipat. Hal ini memungkinkan dimerisasi dan trans-auto-fosforilasi domain sitosol mereka. Aktivasi PERK menghasilkan langsung fosforilasi NRF2 di Ser40, menyebabkan translokasi NRF2 ke inti dan aktivasi gen target. Aktivasi IRE1 menginduksi perekrutan TRAF2 diikuti oleh ASK1 dan fosforilasi dan aktivasi JNK. Karena JNK telah dilaporkan memfosforilasi dan mengaktifkan NRF2, masuk akal untuk berpikir bahwa aktivasi IRE1 akan menyebabkan peningkatan aktivitas NRF2.
Banyak penelitian tentang induksi UPR telah dilakukan dengan penghambat protein glikosilasi tunikamisin. NRF2 tampaknya penting untuk pencegahan kematian sel apoptosis yang diinduksi tunikamisin [31] dan aktivasi dalam kondisi ini didorong oleh degradasi autofagik KEAP1 [51]. Oleh karena itu, pembungkaman ekspresi NRF2 yang dimediasi oleh shRNA dalam sel? TC-6, garis sel insulinoma-sel murine, secara signifikan meningkatkan sitotoksisitas yang diinduksi tunikamisin dan menyebabkan peningkatan ekspresi penanda stres ER pro-apoptosis CHOP10. Di sisi lain, aktivasi NRF2 oleh 1,2-dithiole-3-thione (D3T) mengurangi sitotoksisitas tunikamisin dan melemahkan ekspresi CHOP10 dan PERK [52]. Menariknya, neuron penciuman yang diajukan ke aplikasi sistemik tunikamisin meningkatkan NRF2 secara paralel dengan anggota UPR lainnya seperti CHOP, BIP, XBP1 [53]. Hasil ini telah diperluas ke studi in vivo, karena infus ventrikel lateral tunikamisin pada tikus menginduksi ekspresi PERK dan NRF2 di hipokampus disertai dengan defisit kognitif yang signifikan, peningkatan fosforilasi TAU dan deposit A? 42 [54].
NRF2 Mengatur-Up Gen-Gen Utama untuk Pemeliharaan Fisiologi ER
Lumen ER membutuhkan pasokan GSH yang melimpah dari sitosol untuk mempertahankan kimia disulfida. NRF2 memodulasi enzim penting dari metabolisme GSH di otak, seperti transpor sistin / glutamat,? -Glutamat sistein sintetase (? -GS), glutamat-sistein ligase katalitik dan subunit modulator (GCLC dan GCLM), glutathione reduktase (GR) dan glutathione peroksidase (GPX) (ditinjau dalam [55]). Relevansi NRF2 dalam pemeliharaan GSH di UGD didukung oleh temuan bahwa aktivasi farmakologis atau genetik dari NRF2 menghasilkan peningkatan sintesis GSH melalui GCLC / GCLM, sekaligus menghambat ekspresi enzim tersebut dengan NRF2-knockdown yang menyebabkan akumulasi kerusakan. protein dalam ER yang mengarah ke aktivasi UPR [56].
Pada C. elegans beberapa komponen gen target UPR diatur oleh SKN-1, termasuk Ire1, Xbp1 dan Atf6. Meskipun NRF2 mengatur ekspresi beberapa gen peroksidase (PRX) dan glutathione peroksidase (GPX) pada mamalia (ditinjau dalam [57]), hanya GPX8 yang merupakan enzim ER terlokalisasi yang bonafid, menyimpan sinyal pengambilan KDEL [58]. Kehilangan GPX8 menyebabkan aktivasi UPR, kebocoran ERO1 yang membawa hidrogen peroksida ke sitosol dan kematian sel. Hidrogen peroksida berasal dari ERO1? aktivitas tidak dapat berdifusi dari ER ke sitosol karena aksi bersama GPX8 dan PRX4 [59]. Dalam hal ini, analisis array ekspresi gen jalur pertahanan antioksidan menggunakan RNA dari jaringan tikus wild type dan NRF2-null, mengungkapkan bahwa ekspresi GPX8 diturunkan regulasi tanpa adanya NRF2 [60]. Sejalan dengan ini, analisis transkrip dari sampel pasien yang menderita neoplasma myeloproliferative, polycythemia atau myelofibrosis, penyakit juga terkait dengan stres oksidatif dan peradangan kronis tingkat rendah, menunjukkan tingkat ekspresi yang lebih rendah dari NRF2 dan GPX8 dibandingkan dengan subjek kontrol [61]. Belum ada penelitian yang secara khusus melibatkan GPX8 dalam perlindungan otak manusia tetapi analisis transkriptom pada tikus menunjukkan peningkatan GPX8 kompensasi dalam menanggapi MPTP toksin Parkinsonian [62].
Dampak NRF2 pada Disregulasi UPR pada Penyakit Neurodegeneratif
Malfungsi enzim PDI dan aktivasi kronis dari UPR mungkin selanjutnya memulai atau mempercepat neurodegeneration. Neuron yang terkena penyakit, model hewan dari penyakit neurodegeneratif serta jaringan manusia post-mortem dibuktikan dengan pengaturan beberapa penanda UPR di sebagian besar gangguan ini. Perubahan jalur PDI / UPR pada penyakit neurodegeneratif telah ditinjau dengan baik di [63] tetapi sorotan berikut dari sampel post-mortem otak harus dipertimbangkan. Tingkat PDI meningkat pada neuron kusut dan pada pasien Lewy Bodies of AD dan PD, masing-masing [64], [65]. PDI dan ERP57 diregulasi di CSF dari pasien ALS dan otak dari subjek CJD [66], [67], [68]. BIP, PERK, IRE1 dan ATF6 meningkat dalam sampel dari pasien dengan AD, PD atau ALS [69], [70], [71], [67]. BIP, CHOP dan XBP1 meningkat pada sampel otak post-mortem dari HD [72], [73]. Selain itu, up-regulasi ERP57, GRP94 dan BIP ditemukan dalam jaringan korteks dari pasien CJD [74]. Secara keseluruhan, bukti ini mengungkapkan bahwa akumulasi dari protein yang gagal melipat dalam parenkim otak mengarah pada aktivasi UPR yang merusak dan kronis. Yang menarik, ada penelitian terbaru yang mengaitkan aktivasi NRF2 oleh PERK pada awal AD. Dalam studi ini, penulis menganalisa apakah stres oksidatif yang memediasi perubahan pada NRF2 dan UPR dapat merupakan kejadian awal dalam patogenesis AD dengan menggunakan sel darah perifer manusia dan model tikus transgenik AD pada stadium penyakit yang berbeda. Peningkatan stres oksidatif dan peningkatan pSer40-NRF2 diamati pada sel mononuklear darah perifer manusia yang diisolasi dari individu dengan kerusakan kognitif ringan. Selain itu, mereka melaporkan kerusakan homeostasis kalsium ER dan penanda ER-stres yang diatur di dalam sel-sel ini dari individu dengan kerusakan kognitif ringan dan AD ringan [75].
Regulasi Bersama NRF2 dan Ubiquitin Proteasome�System (UPS)
UPS memodulasi tingkat protein NRF2
UPS berpartisipasi dalam degradasi protein yang rusak atau gagal melipat dan mengontrol tingkat molekul pengatur utama dalam sitosol dan nukleus. Inti inti dari sistem ini adalah enzim multisubunit besar yang mengandung kompleks aktif proteolitik bernama 20S. Proteinome inti 20S mendegradasi protein yang tidak dilipat, tetapi mengikat ke kompleks protein pengatur yang berbeda mengubah spesifisitas substrat dan aktivitasnya. Sebagai contoh, penambahan satu atau dua subunit peraturan 19S ke inti 20S merupakan proteasome 26S dan mengubah spesifisitasnya terhadap protein terlipat asli [76], [77]. Degradasi protein membutuhkan pengikatan kovalen ubiquitin. Konjugasi hasil ubiquitin melalui mekanisme kaskade tiga langkah. Pertama, enzim pengaktif ubiquitin E1 mengaktifkan ubiquitin dalam reaksi yang membutuhkan ATP. Kemudian, satu enzim E2 (protein pembawa ubiquitin atau enzim konjugasi ubiquitin) memindahkan ubiquitin yang diaktifkan dari E1 ke substrat yang secara khusus terikat pada anggota famili ligase ubiquitin-protein, bernama E3. Meskipun nasib pasti dari ubiquitinated-protein akan tergantung pada sifat rantai ubiquitin, proses ini umumnya menghasilkan degradasi oleh 26S proteasome [78].
E3-ligase KEAP1 adalah inhibitor NRF2 yang paling dikenal. Mekanisme regulasi KEAP1 secara elegan menjelaskan bagaimana tingkat NRF2 menyesuaikan dengan fluktuasi oksidan. Dalam kondisi basal, NRF2 yang baru disintesis diambil oleh KEAP1 homodimer, yang mengikat satu molekul NRF2 pada dua sekuens asam amino dengan rendah (aspartat, leusin, glisin, DLG) dan afinitas tinggi (glutamat, treonin, glisin, glutamat, ETGE). Interaksi dengan KEAP1 membantu menyajikan NRF2 ke kompleks protein CULLIN3 / RBX1, yang menyebabkan ubiquitinasi dan degradasi proteasomal berikutnya. Namun, modifikasi redox KEAP1 menghambat penyajian NRF2 ke UPS yang diwakili oleh CULLIN3 / RBX1. Akibatnya, NRF2 yang baru saja disintesis lolos degradasi bergantung-bergantung KEAP1, terakumulasi dalam nukleus dan mengaktifkan gen-gen yang mengandung AD [79], [80], [81], [82].
Adaptor E3-ligase? -TrCP juga merupakan homodimer yang berpartisipasi dalam peristiwa pensinyalan yang terkait dengan fosforilasi NRF2 oleh GSK-3 ?. Kinase ini memfosforilasi residu serin spesifik NRF2 (aspartat, serin, glisin, isoleusin serin; DSGIS) untuk membuat domain degradasi yang kemudian dikenali oleh? -TrCP dan ditandai untuk degradasi proteasome oleh kompleks CULLIN1 / RBX1. Identifikasi asam amino spesifik yang difosforilasi oleh GSK-3? dalam degron ini dilakukan dengan kombinasi mutagenesis langsung dari domain Neh6, elektroforesis 2D-gel [15], [26] dan spektroskopi massa [83]. Akibatnya, penghambatan GSK-3? oleh obat yang sangat selektif atau siRNA terhadap isoform GSK-3 mengakibatkan peningkatan kadar protein NRF2. Hasil serupa ditemukan dengan siRNA terhadap? -TrCP isoform 1 dan 2. Stabilisasi NRF2 mengikuti GSK-3? Penghambatan terjadi pada fibroblas embrio tikus yang kekurangan KEAP1 dan pada mutan penghapusan NRF2 yang diekspresikan secara ektopik yang tidak memiliki residu ETGE penting untuk ikatan afinitas tinggi ke KEAP1, selanjutnya menunjukkan regulasi independen KEAP1.
Dalam konteks penyakit neurodegeneratif, kita dapat membayangkan modulasi NRF2 oleh UPS dalam dua cara berbeda. Di satu sisi, sistem KEAP1 akan merasakan ketidakseimbangan redoks yang berasal dari akumulasi protein yang salah lipatan, sementara sumbu GSK-3 /? - TrCP akan bertindak sebagai peserta aktif dalam transduksi sinyal yang diubah oleh hilangnya proteostasis (Gbr. 2).
Gambar 2 UPS secara ketat mengontrol level NRF2. Dalam kondisi homeostatis, tingkat NRF2 yang rendah dipertahankan oleh aksi adaptor ligase E3 KEAP1 dan? -TrCP. Kiri, NRF2 mengikat domain Kelch dari homodimer KEAP1 melalui motif afinitas rendah (DLG) dan tinggi (ETGE). Melalui domain BTB, KEAP1 secara bersamaan mengikat kompleks CULLIN3 / RBX1, memungkinkan NRF2 di mana-mana dan terdegradasi oleh proteasom 26 S. Apalagi GSK-3? fosforilasi residu Ser335 dan Ser338 dari NRF2 untuk membuat domain degradasi (DpSGIpSL) yang kemudian dikenali oleh adaptor ligase ubiquitin? -TrCP dan ditandai untuk degradasi proteasome oleh kompleks CULLIN3 / RBX1. Benar, Setelah terpapar spesies oksigen reaktif atau elektrofil, residu Cys kritis di KEAP1 dimodifikasi, membuat KEAP1 tidak dapat berinteraksi secara efisien dengan NRF2 atau CULLIN3 / RBX1 dan kemudian faktor transkripsi ini meningkatkan waktu paruh dan aktivitas transkripsi menuju gen ARE. Jalur pensinyalan yang mengakibatkan penghambatan GSK-3 ?, seperti fosforilasi AKT di Ser9, mengakibatkan degradasi NRF2 yang terganggu oleh proteasom, akumulasi dan induksi gen target.
NRF2 Meningkatkan Aktivitas UPS Melalui Kontrol Transkripsi dari Subunit Proteasome
NRF2 mengatur ekspresi beberapa subunit proteasom, sehingga melindungi sel dari akumulasi protein beracun. Dua puluh gen terkait proteasome dan ubiquitination tampaknya diatur oleh NRF2, menurut analisis microarray yang luas dari RNA hati yang dibuat dengan D2T penginduksi NRF3 [84]. Dalam penelitian posterior, penulis yang sama membuktikan bahwa ekspresi sebagian besar subunit proteasom 26S meningkat hingga tiga kali lipat pada hati dari tikus yang diobati dengan D3T. Kadar protein subunit dan aktivitas proteasom meningkat secara terkoordinasi. Namun, tidak ada induksi yang terlihat pada tikus di mana faktor transkripsi NRF2 terganggu. Aktivitas promotor subunit proteasom PSMB5 (20S) meningkat dengan ekspresi berlebih NRF2 atau pengobatan dengan aktivator pada fibroblas embrionik tikus, dan ARE diidentifikasi pada promotor proksimal PSMB5 [85]. Aktivasi farmakologis NRF2 menghasilkan peningkatan tingkat ekspresi subunit proteasome perwakilan (PSMA3, PSMA6, PSMB1 dan PSMB5) hanya pada fibroblas manusia non-senescent yang mengandung NRF2 fungsional [86]. Aktivasi NRF2 selama adaptasi terhadap hasil stres oksidatif dalam ekspresi tinggi PSMB1 (20S) dan PA28? subunit (atau S11, pengatur proteasome) [87]. Selain itu, hasil dari sel induk embrionik manusia mengungkapkan bahwa NRF2 mengontrol ekspresi proteasome maturation protein (POMP), sebuah proteasome pendamping, yang pada gilirannya memodulasi proliferasi sel induk embrionik manusia yang memperbaharui diri, tiga diferensiasi lapisan germinal dan pemrograman ulang seluler [ 88]. Secara keseluruhan, studi ini menunjukkan bahwa NRF2 mengatur ekspresi komponen kunci UPS dan oleh karena itu secara aktif berkontribusi pada pembersihan protein yang jika tidak maka akan menjadi racun.
The NRF2-UPS Axis di Neurodegenerative Diseases
Peran UPS dalam penyakit neurodegeneratif adalah bidang perdebatan intensif. Studi awal melaporkan penurunan aktivitas proteasome pada necropsi manusia dari pasien yang terkena beberapa penyakit neurodegeneratif. Namun, penelitian lain yang menggunakan pendekatan in vitro dan in vivo menemukan aktivitas proteasome yang tidak berubah atau bahkan meningkat (ditinjau dalam [89]). Satu penjelasan yang mungkin untuk perbedaan ini adalah bahwa tingkat komponen UPS mungkin berubah selama perkembangan penyakit dan di daerah otak yang berbeda seperti yang telah disarankan untuk target NRF2.
Terlepas dari kontroversi ini, perlu dicatat bahwa up-regulasi gen proteasome yang mengandung ARE akan memperkuat UPS dengan meningkatkan pembersihan protein beracun di otak. Memang, ablasi NRF1, juga modulator dari respon antioksidan, dalam sel-sel saraf menyebabkan gangguan aktivitas proteasome dan neurodegeneration. Percobaan kromatin immunoprecipitation dan analisis transkripsi menunjukkan bahwa PSMB6 diatur oleh NRF1. Selain itu, profil ekspresi gen menyebabkan identifikasi NRF1 sebagai pengatur transkripsi utama gen proteasome dalam neuron, menunjukkan bahwa gangguan pada NRF1 dapat berkontribusi pada patogenesis penyakit neurodegeneratif [90]. Menariknya, NRF1 dan isoform panjangnya yang disebut TCF11 ditunjukkan untuk meningkatkan gen proteasome yang mengandung AD pada penghambatan proteasome dalam umpan balik untuk mengkompensasi aktivitas proteolitik yang berkurang [91], [92].
Mengenai NRF2, ada korelasi antara pengurangan NRF2, RPT6 (19 S) dan tingkat PSMB5 (20 S) di otak tengah tikus defisien DJ-1 yang diterapi dengan neurotoxin paraquat [93]. Selain itu, senyawa sulforaphane (SFN) yang terjadi secara alami memberikan citra NRF2 yang lebih kuat sebagai modulator penting dari UPS. Percobaan in vitro dengan neuroblastoma murine Sel Neuro2A membuktikan ekspresi yang ditingkatkan dari subunit katalitik proteasome, serta aktivitas peptidase dalam menanggapi SFN. Obat ini melindungi sel dari sitotoksisitas yang dimediasi hidrogen peroksida dan oksidasi protein dengan cara yang bergantung pada fungsi proteasome [94]. Selain itu, Liu dan rekan kerja menggunakan mouse reporter untuk memantau aktivitas UPS dalam menanggapi SFN di otak. Tikus-tikus ini di mana-mana mengekspresikan protein fluoresensi hijau (GFP) menyatu dengan sinyal degradasi konstitutif yang mendorong degradasi cepat oleh UPS (GFPu). Dalam korteks serebral, SFN mengurangi tingkat GFPu dengan peningkatan paralel dalam aktivitas seperti chymotrypsin (PSMB5), caspase-like (PSMB2), dan trypsin (PSMB1) dari proteasome 20. Selain itu, pengobatan sel-sel yang diturunkan Huntington dengan SFN mengungkapkan bahwa aktivasi NRF2 meningkatkan degradasi mHtt dan mengurangi sitotoksisitas mHtt [95]. Mekanisme utama tindakan SFN adalah melalui induksi NRF2 [96]. Kontribusi spesifik NRF2 harus ditangani menggunakan sistem NRF2-null dalam studi lebih lanjut.
Koneksi Fungsional Antara NRF2 dan Macroautophagy
Tingkat Protein NRF2 Dimodulasi oleh Adaptor Protein P62
Autophagy mengacu pada degradasi komponen sitosol di dalam lisosom. Proses ini digunakan untuk pembersihan protein yang berumur panjang dan salah lipat serta organel yang rusak. Sebuah hubungan langsung antara NRF2 dan autophagy pertama kali diamati sehubungan dengan protein adaptor p62, juga disebut SQSTM1 [97], [98], [99], [100], [101]. Protein ini mengangkut protein ubiquitinated ke mesin degradasi proteasomal dan lisosom dan sekuester merusak protein menjadi agregat sebelum degradasi. P62 menyajikan domain ubiquitin-associated (UBA), untuk mengikat ke ubiquitinated proteins, dan sebuah region yang berinteraksi LC3 (LIR) untuk integrasi dengan membran autophagosomal melalui autophagy receptor LC3.
Meskipun induksi NRF62 yang dimediasi p2 dan gen targetnya pertama kali dilaporkan pada tahun 2007 [102], mekanisme molekuler tidak sepenuhnya dipahami sampai ditemukan interaksinya dengan KEAP1 [103], [98], [99], [100 ], [101]. Komatsu dan rekan kerjanya mengidentifikasi wilayah interaksi (KIR) KEAP1 di p62 yang mengikat KEAP1 di kantong permukaan dasar yang sama dengan NRF2 dan dengan afinitas pengikatan yang mirip dengan motif ETGE di NRF2, menunjukkan persaingan antara p62 dan NRF2. Fosforilasi Ser351 dalam motif KIR di p62 (349-DPSTGE-354) terbukti meningkatkan afinitasnya untuk KEAP1, bersaing dengan pengikatan NRF2 dan memungkinkan akumulasi dan aktivasi transkripsi dari gen targetnya [98], [99]. Faktanya, ekspresi berlebih p62 menyebabkan berkurangnya NRF2 di mana-mana dan akibatnya stabilisasi serta induksi gen targetnya [104]. Beberapa kinase telah disarankan untuk berpartisipasi dalam fosforilasi p62. Target mamalia dari rapamycin complex 1 (mTORC1) mungkin terlibat, karena pengobatan dengan mTOR inhibitor rapamycin menekan fosforilasi p62 dan penurunan regulasi KEAP1 pada pengobatan arsenit. Baru-baru ini, ditunjukkan bahwa TGF -? - kinase 1 yang diaktifkan (TAK1) juga dapat memfosforilasi p62, meningkatkan degradasi KEAP1 dan regulasi peningkatan NRF2. Penulis penelitian ini menyarankan ini adalah cara untuk mengatur redokstasis seluler dalam kondisi mapan, karena defisiensi TAK1 mengatur ROS dengan tidak adanya oksidan eksogen di jaringan tikus yang berbeda secara paralel dengan penurunan kadar protein NRF2 [105 ].
Konstruksi p62 yang tidak memiliki domain UBA masih mampu mengikat KEAP1, menyiratkan bahwa interaksi tidak bergantung pada KEAP1 [101] yang mana-mana. Namun, homolog p62 di Drosophila melanogaster, bernama Ref (2), tidak mengandung motif KIR dan tidak secara langsung berinteraksi dengan DmKEAP1, meskipun dapat mengikat DmKEAP1 yang dimanisakan melalui domain UBA. Selain itu, DmKEAP1 dapat langsung berinteraksi dengan Atg8 (homolog ke mamalia LC3). KEAP1 kekurangan hasil di Atg8 dan induksi autophagy tergantung pada NRF2 orthologue CncC dan independen pada TFEB / MITF [106]. Hubungan antara NRF2 dan autophagy tampaknya dilestarikan meskipun, menyoroti relevansi fungsionalnya.
Induksi NRF2 oleh p62 adalah hasil dari kedua kompetisi untuk mengikat KEAP1 dan degradasi KEAP1 di lisosom. Membungkam p62 dengan siRNA menggandakan KEAP1 paruh waktu secara paralel dengan penurunan NRF2 dan gen targetnya [101]. Dalam kesepakatan, ablasi ekspresi p62 dibuktikan peningkatan tingkat KEAP1 dibandingkan dengan tikus wild type. Sangat relevan, peningkatan dalam tingkat KEAP1 tidak dipengaruhi oleh inhibitor proteasome tetapi berkurang di bawah autophagy yang menyebabkan kelaparan [107]. Bahkan, KEAP1 hadir dalam sel mamalia di vesikel autophagic yang dihiasi dengan p62 dan LC3 [99], [100], [103]. Semua data ini menunjukkan bahwa KEAP1 adalah substrat dari mesin macroautophagy, tetapi masalah ini harus dianalisis dengan lebih detail karena adanya beberapa hasil yang kontroversial. Kadar protein KEAP1 meningkat pada tikus Atg7-null, efektor kunci dari macroautophagy [107], tetapi inhibisi farmakologi dari macroautophagy dengan torin1, E64 / pepstatin atau bafilomycin gagal mengakumulasi KEAP1 [107], [100]. Secara keseluruhan, hasil ini menunjukkan bahwa peningkatan tingkat p62 menyita KEAP1 menjadi vakuola autophagic dan mungkin hasil ini dalam degradasi autophagic KEAP1 memungkinkan aktivasi NRF2 (Gambar 3). Dua penelitian yang berbeda melaporkan bahwa asam sulfinat reduktase SESTRIN memainkan peran penting dalam konteks ini. SESTRIN 2 berinteraksi dengan p62, KEAP1, dan RBX1 serta memfasilitasi degradasi dependen P62 dari KEAP1 dan NRF2 dari gen target [108]. Studi lain menunjukkan bahwa SESTRIN 2 berinteraksi dengan ULK1 dan p62, mempromosikan fosforilasi p62 di Ser403 yang memfasilitasi degradasi protein kargo termasuk KEAP1 [109].
Gambar tingkat 3 NRF2 diatur oleh protein adaptor p62. The fosforilasi Ser 351 dalam motif KIR dari p62 (349-DPSTGE-354) oleh mTORC1, TAK1 atau kinase lainnya menghasilkan peningkatan afinitas untuk mengikat KEAP1 karena kemiripan dengan motif ETGE di NRF2. Sebagai akibatnya, p62 terfosforilasi menggantikan NRF2 dan mengikat KEAP1. Motif LIR di p62 memungkinkan interaksi dengan LC3 di membran autophagosomal, sehingga kompleks p62-KEAP1 akhirnya terdegradasi di lisosom. Sebagai akibatnya NRF2 mampu berakumulasi, mentranslokasi ke nukleus dan meningkatkan transkripsi gen yang mengandung ARE, termasuk p62. Mekanisme pengaturan ini memberikan respons NRF2 yang dapat diperdebatkan, karena KEAP1 harus disintesis baru untuk menghambat aktivitas NRF2.
Modulasi Gen Macroautophagy oleh NRF2
NRF2 mengatur ekspresi gen yang relevan untuk macroautophagy dan juga untuk UPR dan UPS. Bukti pertama berasal dari penelitian di mana ekspresi p62 terbukti diinduksi pada paparan elektrofil, ROS dan nitrat oksida [110], [111], [112]. Mekanisme induksi dijelaskan beberapa tahun kemudian dengan temuan bahwa p62 berisi ARE fungsional dalam promotor gennya [99]. Dalam penelitian terbaru, beberapa ARE fungsional lainnya ditemukan dan divalidasi berikut analisis bioinformatika dan tes ChIP. Selain itu, fibroblas embrio tikus dan neuron kortikal dari tikus Nrf2-knockout menunjukkan penurunan ekspresi p62, yang dapat diselamatkan dengan lisisvirus NRF2-expressing. Demikian pula, defisiensi NRF2 mengurangi tingkat p62 pada neuron yang cedera dari tikus hippocampus [36]. Oleh karena itu, telah disarankan bahwa aktivasi NRF2 meningkatkan level p62, menghasilkan degradasi KEAP1 dan mendukung stabilisasi NRF2 lebih lanjut dalam loop umpan balik positif. Mekanisme non-kanonik ini dari induksi NRF2 membutuhkan perubahan dalam ekspresi gen dan mungkin merupakan respons yang relevan terhadap stres seluler yang berkepanjangan.
Protein pengenalan muatan NDP52 ditunjukkan diatur secara transkripsi oleh NRF2. NDP52 bekerja dengan cara yang mirip dengan p62, mengenali protein ubiquitinated dan berinteraksi dengan LC3 melalui domain LIR, sehingga kargo terdegradasi di lisosom. Lima Warisan yang diduga ditemukan dalam urutan DNA promoter Ndp52. Tiga dari mereka diidentifikasi dengan konstruksi mutan yang berbeda dan tes CHIP sebagai sangat diperlukan untuk transkripsi Ndp2 yang dimediasi NRF52 [113]. Dari catatan, tingkat mRNA Ndp52 berkurang pada hippocampus tikus Nrf2-knockout. Salah satu dari sekuens ini juga divalidasi dalam penelitian independen sebagai ARE yang diatur oleh NRF2 [36].
Namun, peran NRF2 dalam modulasi autophagy tidak terbatas pada induksi dua protein pengenal muatan ini. Untuk mendapatkan wawasan yang lebih dalam tentang peran NRF2 dalam modulasi gen terkait autophagy tambahan, kelompok kami menskrining basis data kromatin imunopresipitasi ENCODE untuk dua protein, MAFK dan BACH1, yang mengikat NRF2 yang diatur oleh ARE. Dengan menggunakan skrip yang dihasilkan dari urutan konsensus JASPAR, kami mengidentifikasi beberapa ARE yang diduga dalam banyak gen autophagy. Dua belas dari sekuens ini divalidasi sebagai NRF2 yang mengatur AREs dalam sembilan gen autophagy, yang ekspresinya berkurang pada fibroblast embrio tikus dari Nrf2-knockout tikus tetapi dapat dipulihkan oleh lentivirus yang mengekspresikan NRF2. Studi kami menunjukkan bahwa NRF2 mengaktifkan ekspresi beberapa gen yang terlibat dalam langkah-langkah berbeda dari proses autophagic, termasuk inisiasi autophagy (ULK1), pengenalan kargo (p62 dan NDP52), pembentukan autophagosome (ATG4D, ATG7 dan GABX1XXXXX2XXXXUMUM5). ), dan izin autolysosome (ATG4D). Akibatnya, fluks autofagi dalam menanggapi hidrogen peroksida terganggu ketika NRF2 tidak ada [36].
Relevansi Ekspresi Otak Makroautofagi NRFPNUMX-Mediated di Neurodegenerative Disorders
Autophagy yang rusak telah terbukti memainkan peran penting dalam beberapa penyakit neurodegeneratif [114] dan ablasi autophagy menyebabkan neurodegeneration pada tikus [115], [116]. Tikus Atg7-knockout mengungkapkan bahwa defisiensi autophagy menghasilkan akumulasi p62 di badan inklusi ubiquitin-positif. KEAP1 diasingkan di badan inklusi ini, mengarah ke stabilisasi NRF2 dan induksi gen target [103]. Yang penting, akumulasi berlebihan p62 bersama dengan protein ubiquitinated telah diidentifikasi dalam penyakit neurodegeneratif, termasuk AD, PD dan ALS [117]. Bahkan, neuron yang mengekspresikan tingkat tinggi APP atau TAU pasien AD juga menyatakan p62 dan NRF2 nuklir, menunjukkan upaya mereka untuk menurunkan agregat intraneuronal melalui autophagy [36].
Kekurangan NRF2 memperburuk agregasi protein dalam konteks DA. Faktanya, peningkatan level TAU terfosforilasi dan tidak larut sarkosil ditemukan pada tikus Nrf2-knockout, meskipun tidak ada perbedaan dalam aktivitas kinase atau fosfatase yang dapat dideteksi dibandingkan dengan latar belakang tipe liar [113]. Yang penting, NDP52 didemonstrasikan untuk bekerja sama dengan TAU di neuron murine dan interaksi langsung antara fosfo-TAU dan NDP52 ditunjukkan oleh percobaan co-immunoprecipitation baik pada tikus dan sampel AD, menunjukkan perannya dalam degradasi TAU. Menariknya, pembungkaman NDP52, p62 atau NRF2 di neuron menghasilkan peningkatan fosfo-TAU [113], [118]. Selain itu, peningkatan agregat APP intraneuronal ditemukan di hipokampus mencit APP / PS1? E9 ketika NRF2 tidak ada. Ini berkorelasi dengan penanda autofagi yang diubah, termasuk peningkatan rasio fosfo-mTOR / mTOR dan fosfo-p70S6k / p70S6k (indikasi penghambatan autofagi), peningkatan kadar pra-katepsin D dan sejumlah besar badan multivesikuler [119]. Pada tikus yang ikut mengekspresikan APP manusia (V717I) dan TAU (P301L), defisiensi NRF2 menyebabkan peningkatan kadar total dan fosfo-TAU dalam fraksi yang tidak larut dan peningkatan agregat APP intraneuronal, bersama dengan penurunan kadar neuronal p62, NDP52, ULK1, ATG5 dan GABARAPL1. Ko-lokalisasi antara protein adaptor p62 dan APP atau TAU berkurang dengan tidak adanya NRF2 [36]. Secara keseluruhan, hasil ini menyoroti pentingnya NRF2 dalam autofagi neuronal.
Berbagai Faktor Transkripsi Bertindak Koordinasi untuk Memodulasi Proteostasis
Di bawah kondisi steady state, proteostasis dikendalikan melalui interaksi protein-protein dan modifikasi pasca-translasi mendapatkan respon cepat. Namun, adaptasi seluler membutuhkan pengaturan transkripsi gen UPR, UPS dan autophagy. Mempertimbangkan bahwa sel-sel saraf secara terus-menerus diserahkan kepada penghilang racun tingkat rendah, termasuk stres oksidatif dan proteotoksik, penguatan proteostasis yang disebabkan oleh modulasi transkripsi dapat membantu mencegah degenerasi otak.
Dalam kasus UPR, aktivasi masing-masing dari ketiga lengan akhirnya akan menghasilkan induksi transkripsi gen tertentu (ditinjau dalam [43]). Sebagai contoh, sebuah fragmen ATF6 yang diturunkan (ATF6f) berikatan dengan elemen-elemen respons ER-stress (ERSE) dan menginduksi ekspresi beberapa gen, termasuk XBPI, BIP dan CHOP. Selain itu, pensinyalan PERK mengarah pada aktivasi faktor transkripsi ATF4, yang mengontrol ekspresi beberapa gen terkait UPR dan beberapa lainnya termasuk gen target NRF2 Hmox1 dan p62. Akhirnya, aktivasi IRE1 menghasilkan generasi faktor transkripsi aktif, XBP1 yang disambung (XBP1s), yang mengontrol transkripsi gen yang mengkode protein yang terlibat dalam pelipatan protein.
Di sisi lain, NRF1 terbukti diperlukan untuk ekspresi gen proteasomal di otak, karena tikus Nrf1-knockout menunjukkan berkurangnya ekspresi gen yang mengkode berbagai subunit inti 20S, serta kompleks pengaturan 19S bersama dengan fungsi proteasomal yang terganggu [90 ]. NRF1 dan NRF2 terikat pada urutan ARE di wilayah promotor gen targetnya, yang menunjukkan bahwa mereka memiliki aktivitas transkrip yang tumpang tindih, meskipun mereka berbeda dalam mekanisme pengaturan dan lokalisasi seluler [120].
Faktor-faktor transkripsi dari keluarga kotak Forkhead O (FOXO) mengendalikan ekspresi gen-gen yang berhubungan dengan autophagy. Mirip dengan apa yang terjadi dengan NRF2, ada beberapa lapisan regulasi aktivitas anggota FOXO, yang dapat diinduksi pada stres nutrisi atau oksidatif [121]. Akhirnya, faktor transkripsi TFEB, dianggap sebagai pengatur utama dari biogenesis lisosom, memainkan peran penting dalam pengaturan autophagy di bawah kondisi stres nutrisi. Dengan demikian, penghambatan mTORC1 menyebabkan translokasi nuklir TFEB dan induksi ekspresi gen autophagy [122].
Secara keseluruhan, keberadaan regulator transkripsi yang berbeda dari mesin-mesin ini juga menunjukkan crosstalk dan mekanisme sebagian redundan yang dapat menjamin proteostasis dalam keadaan yang berbeda. Dengan demikian, NRF2 mungkin memiliki peran yang relevan dalam jaringan yang mendukung tingkat stres oksidatif yang tinggi. Misalnya, NRF2 yang diinduksi stres oksidatif mungkin berfungsi dalam kondisi yang kaya nutrisi secara transkripsi hingga mengatur autophagy, mirip dengan apa yang telah ditemukan untuk TFEB dalam kondisi kelaparan. Selain itu, fungsi otak sebagian besar di bawah kondisi kaya nutrisi, berpose NRF2 sebagai mekanisme yang relevan untuk mengaktifkan autophagy di neuron.
Menjanjikan Potensi Terapi untuk NRF2 di Proteinopathies
Dalam beberapa tahun terakhir, kemajuan besar telah dibuat dalam pengetahuan tentang peran pengaturan dari UPR, UPS dan autophagy pada aktivitas NRF2, serta rekam jejak yang dimediasi NRF2 dari komponen ketiga sistem ini. Oleh karena itu, kemungkinan terapeutik baru mungkin timbul berdasarkan eksploitasi NRF2 sebagai pengatur penting pembebasan protein pada penyakit neurodegeneratif.
Namun, pertanyaan kunci yang tersisa adalah apakah akan berguna atau merusak untuk meningkatkan tingkat NRF2 di otak. Analisis data epidemiologi dapat memberikan jawaban parsial, karena hal ini menunjukkan bahwa gen NFE2L2 sangat polimorfik dan beberapa polimorfisme nukleotida tunggal yang ditemukan di wilayah regulasi promotornya dapat memberikan kisaran variabilitas fisiologis dalam ekspresi gen pada tingkat populasi dan beberapa haplotipe dikaitkan dengan penurunan risiko dan / atau keterlambatan timbulnya DA, PD atau ALS [123]. Selain itu, seperti yang dibahas oleh Hayes dan rekan [124], efek NRF2 mungkin memiliki respons berbentuk U, yang berarti bahwa tingkat NRF2 yang terlalu rendah dapat mengakibatkan hilangnya perlindungan sitoproteksi dan peningkatan kerentanan terhadap stresor, sementara terlalu banyak NRF2 dapat mengganggu keseimbangan homeostatis. skenario reduktif (stres reduktif), yang akan mendukung kesalahan lipatan dan agregasi protein. Tingkat NRF2 yang rendah di otak mendukung gagasan bahwa sedikit peningkatan regulasi mungkin cukup untuk mencapai manfaat dalam kondisi patologis. Faktanya, peran protektif dari aktivasi farmakologis NRF2 yang dimediasi dari pembersihan protein telah ditunjukkan dalam kultur sel neurodegenerasi yang berbeda dan model in vivo.
SFN adalah aktivator NRF2 farmakologis yang ditunjukkan untuk menginduksi ekspresi gen proteasomal dan autophagy [95], [36]. Menariknya, Jo dan rekan menunjukkan bahwa SFN mengurangi tingkat TAU terfosforilasi dan meningkatkan Beclin-1 dan LC3-II, menunjukkan aktivasi NRF2 dapat memfasilitasi degradasi protein beracun ini melalui autophagy [113]. Selain itu, degradasi mHtt ditingkatkan dengan SFN, dan ini dikembalikan dengan penggunaan MG132, menunjukkan degradasi proteasomal dari protein beracun ini [95]. Degradasi fosfo- dan tidak larut-TAU yang dimediasi autofagi dilaporkan dengan fisetin flavonoid organik. Senyawa ini mampu menginduksi autofagi dengan secara bersamaan mempromosikan aktivasi dan translokasi inti dari TFEB dan NRF2, bersama dengan beberapa gen targetnya. Tanggapan ini dicegah dengan pembungkaman TFEB atau NRF2 [125]. Bott dan rekannya melaporkan efek menguntungkan dari aktivator NRF2, NRF1 dan HSF1 simultan pada toksisitas protein pada atrofi otot tulang belakang dan bulbar, gangguan neurodegeneratif yang disebabkan oleh perluasan pengulangan CAG pengkode poliglutamin di mana agregat protein hadir [126]. Potensi aktivasi NRF2 untuk pengobatan gangguan neurodegeneratif telah dibuktikan dengan persetujuan BG-12, formulasi oral dari NRF2 inducer dimethyl fumarate (DMF), untuk pengobatan multiple sclerosis [127], [128]. Keberhasilan DMF dengan penyakit autoimun dengan komponen inflamasi yang kuat menunjukkan bahwa penyakit neurodegeneratif mungkin mendapat manfaat dari reposisi obat ini. Dalam studi praklinis baru-baru ini dari model? -Synucleinopathy dari PD, DMF terbukti menjadi pelindung saraf karena, sebagian, untuk induksi autofagi [129]. Studi yang melaporkan efek menguntungkan dari NRF2 pada neurodegenerasi tetapi tidak berfokus pada efeknya pada pembersihan protein bahkan lebih berlimpah (untuk tinjauan komprehensif, lihat [7]). Ini cukup relevan, karena menyoroti beberapa proses merusak yang secara bersamaan dapat ditargetkan oleh satu pukulan di NRF2, juga termasuk stres oksidatif, peradangan saraf, atau disfungsi mitokondria. Namun, pekerjaan masa depan akan diperlukan untuk menentukan secara pasti apakah aktivasi farmakologis NRF2 mungkin merupakan strategi yang valid untuk memfasilitasi degradasi protein beracun di otak.
Seperti yang dijelaskan sebelumnya, memperburuk GSK-3? aktivitas dilaporkan pada penyakit neurodegeneratif dan telah berspekulasi bahwa akibat pengurangan NRF2 dapat sebagian bertanggung jawab atas hasil yang merusak. Di bawah kondisi patologis ini, penghambat GSK-3 juga dapat bekerja sama untuk meningkatkan tingkat NRF2 dan proteostasis. Efek menguntungkan dari penghambat GSK-3 telah dilaporkan dalam berbagai model neurodegenerasi dan, yang lebih menarik, represi GSK-3 terbukti mengurangi tingkat protein beracun [130], [131], [132], [133]. Meskipun belum ada hubungan langsung antara penghambatan GSK-3 dan regulasi transkripsional NRF2 dari gen yang mempromosikan proteostasis yang telah diamati, masuk akal untuk berspekulasi bahwa penurunan regulasi aktivitas GSK-3 akan menghasilkan peningkatan tingkat NRF2, yang pada akhirnya akan menghasilkan penguatan proteostasis.
Aktivitas transkripsi NRF2 serta kapasitas sel untuk mempertahankan proteostasis menurun seiring bertambahnya usia, faktor risiko utama perkembangan penyakit neurodegeneratif. Masuk akal untuk berpikir bahwa penguatan NRF2 dan, akibatnya, proteostasis akan, setidaknya, menunda akumulasi agregat protein dan neurodegenerasi. Memang, pengobatan fibroblas tua manusia dengan 18? -Glycyrrhetinic acid (18? -GA) triterpenoid mempromosikan aktivasi NRF2, yang mengarah ke induksi proteasome dan meningkatkan masa hidup. Studi ini menunjukkan bahwa aktivasi farmakologis NRF2 dimungkinkan bahkan di usia lanjut [86]. Selain itu, penelitian selanjutnya menunjukkan bahwa senyawa ini memediasi SKN-1 dan aktivasi proteasome di C.elegans dengan efek menguntungkan pada perkembangan DA dalam model nematoda yang relevan [134].
Semua hal dipertimbangkan, induksi NRF2-induksi gen terkait-proteostasis tampaknya bermanfaat dalam berbagai proteinopathies.
Sulforaphane dan Dampaknya pada Kanker, Kematian, Penuaan, Otak dan Perilaku, Penyakit Jantung & Lainnya
Isothiocyanate adalah beberapa senyawa tanaman yang paling penting yang bisa Anda dapatkan dalam diet Anda. Dalam video ini saya membuat kasus yang paling komprehensif untuk mereka yang pernah dibuat. Rentang perhatian yang pendek? Lewati ke topik favorit Anda dengan mengklik salah satu poin waktu di bawah ini. Garis waktu penuh di bawah ini.
Bagian utama:
00: 01: 14 - Kanker dan kematian
00: 19: 04 - Penuaan
00: 26: 30 - Otak dan perilaku
00: 38: 06 - Rekap terakhir
00: 40: 27 - Dosis
Lini waktu penuh:
00: 00: 34 - Pengantar sulforaphane, fokus utama dari video.
00: 01: 14 - konsumsi sayuran Cruciferous dan pengurangan dalam semua penyebab kematian.
00: 02: 12 - Risiko kanker prostat.
00: 02: 23 - Risiko kanker kandung kemih.
00: 02: 34 - Kanker paru-paru berisiko pada perokok.
00: 02: 48 - Risiko kanker payudara.
00: 03: 13 - Hipotetis: bagaimana jika Anda sudah menderita kanker? (intervensi)
00: 03: 35 - Mekanisme yang masuk akal mengemudi kanker dan data asosiatif kematian.
00: 04: 38 - Sulforaphane dan kanker.
00: 05: 32 - Hewan bukti yang menunjukkan efek kuat dari ekstrak kecambah brokoli pada perkembangan tumor kandung kemih pada tikus.
00: 06: 06 - Pengaruh suplementasi langsung sulforaphane pada pasien kanker prostat.
00: 07: 09 - Bioakumulasi metabolit isothiocyanate dalam jaringan payudara yang sebenarnya.
00: 08: 32 - Penghambatan sel induk kanker payudara.
00: 08: 53 - Pelajaran sejarah: brassica ditetapkan memiliki sifat-sifat kesehatan bahkan di Roma kuno.
00: 09: 16 - Kemampuan Sulforaphane untuk meningkatkan ekskresi karsinogen (benzena, akrolein).
00: 09: 51 - NRF2 sebagai saklar genetik melalui elemen respons antioksidan.
00: 10: 10 - Bagaimana aktivasi NRF2 meningkatkan ekskresi karsinogen melalui glutathione-S-conjugates.
00: 10: 34 - kubis Brussel meningkatkan glutathione-S-transferase dan mengurangi kerusakan DNA.
00: 11: 20 - Broccoli sprout drink meningkatkan ekskresi benzena oleh 61%.
00: 13: 31 - Broccoli sprout homogenate meningkatkan enzim antioksidan di saluran napas bagian atas.
00: 15: 45 - konsumsi sayuran Cruciferous dan kematian penyakit jantung.
00: 16: 55 - Bubuk tunas brokoli meningkatkan lipid darah dan risiko penyakit jantung secara keseluruhan pada penderita diabetes tipe 2.
00: 19: 04 - Awal dari bagian penuaan.
00: 19: 21 - Diet yang diperkaya Sulforaphane meningkatkan masa hidup kumbang dari 15 ke 30% (dalam kondisi tertentu).
00: 20: 34 - Pentingnya peradangan rendah untuk umur panjang.
00: 22: 05 - Sayuran dan tunas kecambah brokoli tampaknya mengurangi beragam penanda inflamasi pada manusia.
00: 23: 40 - Rekap video pertengahan: kanker, bagian penuaan
00: 24: 14 - Studi pada tikus menunjukkan sulforaphane dapat meningkatkan fungsi imun adaptif di usia tua.
00: 25: 18 - Sulforaphane meningkatkan pertumbuhan rambut pada model tikus botak. Gambar di 00: 26: 10.
00: 26: 30 - Awal dari bagian otak dan perilaku.
00: 27: 18 - Pengaruh ekstrak kecambah brokoli pada autisme.
00: 27: 48 - Pengaruh glucoraphanin pada skizofrenia.
00: 28: 17 - Mulai dari diskusi depresi (mekanisme dan studi yang masuk akal).
00: 31: 21 - Studi mouse menggunakan 10 model yang berbeda dari depresi yang diinduksi stres menunjukkan sulforaphane sama efektifnya dengan fluoxetine (prozac).
00: 32: 00 - Studi menunjukkan konsumsi langsung glukoraphanin pada tikus juga efektif mencegah depresi dari model stres kekalahan sosial.
00: 33: 01 - Awal dari bagian neurodegenerasi.
00: 33: 30 - Sulforaphane dan penyakit Alzheimer.
00: 33: 44 - Sulforaphane dan penyakit Parkinson.
00: 33: 51 - Sulforaphane dan penyakit Hungtington.
00: 34: 13 - Sulforaphane meningkatkan protein heat shock.
00: 34: 43 - Awal dari bagian cedera otak traumatis.
00: 35: 01 - Sulforaphane disuntikkan segera setelah TBI meningkatkan daya ingat (studi pada tikus).
00: 35: 55 - Sulforaphane dan plastisitas neuronal.
00: 36: 32 - Sulforaphane meningkatkan pembelajaran dalam model diabetes tipe II pada tikus.
00: 37: 19 - Sulforaphane dan duchenne muscular dystrophy.
00: 37: 44 - Myostatin inhibition dalam sel-sel satelit otot (in vitro).
00: 38: 06 - Rekap video-ulang: mortalitas dan kanker, kerusakan DNA, stres oksidatif dan peradangan, ekskresi benzena, penyakit kardiovaskular, diabetes tipe II, efek pada otak (depresi, autisme, skizofrenia, neurodegenerasi), jalur NRF2.
00: 40: 27 - Pikiran tentang mencari tahu kecambah brokoli atau sulforaphane.
00: 41: 01 - Anekdot saat bertumbuh di rumah.
00: 43: 14 - Pada suhu memasak dan aktivitas sulforaphane.
00: 43: 45 - Konversi bakteri usus dari sulforaphane dari glucoraphanin.
00: 44: 24 - Suplemen bekerja lebih baik ketika dikombinasikan dengan myrosinase aktif dari sayuran.
00: 44: 56 - Teknik memasak dan sayuran silangan.
00: 46: 06 - Isothiocyanate sebagai goitrogens.
Faktor nuklir 2 yang diturunkan eritroid (NF-E2) yang berhubungan dengan faktor 2, atau dikenal sebagai Nrf2, adalah faktor transkripsi yang mengatur ekspresi berbagai enzim antioksidan dan detoksifikasi. Studi penelitian juga menunjukkan perannya dalam mengendalikan stres oksidatif. Sebagian besar penyakit neurodegeneratif, seperti penyakit Alzheimer dan Parkinson, ditandai oleh stres oksidatif dan peradangan kronis, target umum dari Pendekatan pengobatan Nrf2. Dr Alex Jimenez DC, CCST Insight
Catatan Penutup
Faktor transkripsi NRF2 mengatur respon proteostatik dengan merasakan dan memodulasi perubahan dalam UPR, UPS dan autophagy (Gambar. 4). Akibatnya, kurangnya NRF2 telah terbukti memperburuk proteinopati, menunjukkan bahwa NRF2 diperlukan untuk pembersihan protein yang optimal. Bersama-sama, kita dapat berspekulasi bahwa NRF2 mungkin menjadi target terapi yang menarik untuk proteinopat.
Gambar 4 NRF2 sebagai hub yang menghubungkan sinyal darurat yang diturunkan dari proteotoxic ke respons transkripsi protektif. Akumulasi protein yang tidak dilipat / gagal melipat akan menyebabkan aktivasi respon protein yang tidak terbuka (UPR) di UGD. Aktivasi PERK atau MAPK dapat menghasilkan induksi transkripsi ER-residen Gpx8 dan beberapa enzim yang mengatur kadar GSH, penting untuk memastikan lipatan protein yang benar. Agregat protein menghambat aktivitas proteasome (UPS), mungkin menghindari degradasi NRF2. NRF2 telah ditunjukkan untuk secara khusus memodulasi transkripsi Psma3, Psma6, Psmb1, Psmb5 dan gen Pomp. Beberapa subunit lainnya diregulasi dengan cara NRF2 sebagai respon terhadap D3T, mungkin memperbesar daftar subunit proteasome yang diatur oleh NRF2. Autophagy adalah jalur utama untuk degradasi agregat protein. Autophagy juga mengatur NRF2, menghubungkan jalur degradasi ini dengan induksi transkripsi NRF2 dari p62, Ndp52, Ulk1, Atg2b, Atg4c, Atg5, Atg7, dan Gabarapl1.
Menurut artikel di atas, sementara gejala penyakit neurodegeneratif dapat diobati melalui berbagai pilihan pengobatan, penelitian telah menunjukkan bahwa aktivasi Nrf2 dapat menjadi pendekatan perawatan yang bermanfaat. Karena Aktivis Nrf2 menargetkan mekanisme luas penyakit, semua penyakit neurodegeneratif dapat memperoleh manfaat dari penggunaan faktor transkripsi Nrf2. Temuan Nrf2 telah merevolusi pengobatan penyakit neurodegeneratif. Cakupan informasi kami terbatas pada masalah chiropraktik dan kesehatan tulang belakang. Untuk membahas pokok bahasan ini, jangan ragu untuk bertanya kepada Dr. Jimenez atau hubungi kami di 915-850-0900 .
Diskusi Topik Tambahan: Menghilangkan Nyeri Lutut tanpa Pembedahan
Nyeri lutut adalah gejala umum yang dapat terjadi karena berbagai cedera dan / atau kondisi lutut, termasuk cedera olahraga. Lutut adalah salah satu sendi yang paling kompleks dalam tubuh manusia karena terdiri dari persimpangan empat tulang, empat ligamen, berbagai tendon, dua menisci, dan tulang rawan. Menurut American Academy of Family Physicians, penyebab paling umum dari nyeri lutut termasuk subluksasi patela, tendinitis patela atau lutut pelompat, dan penyakit Osgood-Schlatter. Meskipun nyeri lutut paling mungkin terjadi pada orang berusia di atas 60 tahun, nyeri lutut juga dapat terjadi pada anak-anak dan remaja. Nyeri lutut dapat diobati di rumah dengan mengikuti metode RICE, namun cedera lutut yang parah mungkin memerlukan perhatian medis segera, termasuk perawatan chiropraktik. �
Stres oksidatif digambarkan sebagai kerusakan sel yang disebabkan oleh radikal bebas, atau molekul tidak stabil, yang pada akhirnya dapat memengaruhi fungsi kesehatan. Tubuh manusia menciptakan radikal bebas untuk menetralisir bakteri dan virus, namun faktor eksternal, seperti oksigen, polusi, dan radiasi, seringkali juga dapat menghasilkan radikal bebas. Stres oksidatif telah dikaitkan dengan berbagai masalah kesehatan.
Stres oksidatif dan stresor lainnya mengaktifkan mekanisme perlindungan internal yang dapat membantu mengatur respons antioksidan tubuh manusia. Nrf2 adalah protein yang merasakan tingkat stres oksidatif dan memungkinkan sel untuk melindungi diri dari faktor internal dan eksternal. Nrf2 juga telah ditunjukkan untuk membantu mengatur gen yang terlibat dalam produksi enzim antioksidan dan gen respons stres. Tujuan artikel di bawah ini adalah untuk menjelaskan efek dari Nrf2 pada kanker.
Abstrak
Jalur Keap1-Nrf2 adalah pengatur utama tanggapan sitoprotektif terhadap stres oksidatif dan elektrofilik. Meskipun jalur signaling sel yang dipicu oleh faktor transkripsi Nrf2 mencegah inisiasi kanker dan perkembangan dalam jaringan normal dan prareligna, pada sel yang sepenuhnya ganas, aktivitas Nrf2 memberikan keuntungan pertumbuhan dengan meningkatkan kemoresisten kanker dan meningkatkan pertumbuhan sel tumor. Dalam ulasan grafis ini, kami memberikan gambaran tentang jalur Keap1-Nrf2 dan disregulasi dalam sel kanker. Kami juga secara singkat merangkum konsekuensi dari aktivasi Nrf2 konstitutif dalam sel kanker dan bagaimana ini dapat dimanfaatkan dalam terapi gen kanker.
Kata kunci:Nrf2, Keap1, Kanker, Unsur respon antioksidan, terapi gen
Pengantar
Jalur Keap1-Nrf2 adalah pengatur utama respons sitoprotektif terhadap tekanan endogen dan eksogen yang disebabkan oleh spesies oksigen reaktif (ROS) dan elektrofil [1]. Protein pensinyalan kunci dalam jalur tersebut adalah faktor transkripsi Nrf2 (faktor nuklir terkait eritroid 2 faktor 2) yang mengikat bersama dengan protein Maf kecil ke elemen respons antioksidan (ARE) di wilayah regulasi gen target, dan Keap1 (Kelch ECH protein asosiasi 1), protein penekan yang mengikat Nrf2 dan mendorong degradasinya oleh jalur proteasome ubiquitin (Gbr. 1). Keap1 adalah protein yang sangat kaya sistein, Keap1 tikus memiliki total 25 dan 27 residu sistein manusia, yang sebagian besar dapat dimodifikasi secara in vitro oleh oksidan dan elektrofil yang berbeda [2]. Tiga dari residu ini, C151, C273 dan C288, telah terbukti memainkan peran fungsional dengan mengubah konformasi Keap1 yang mengarah ke translokasi nuklir Nrf2 dan ekspresi gen target berikutnya [3] (Gbr. 1). Mekanisme yang tepat dimana modifikasi sistein di Keap1 menyebabkan aktivasi Nrf2 tidak diketahui, tetapi dua model yang berlaku tetapi tidak saling eksklusif adalah (1) model 'engsel dan kait', di mana modifikasi Keap1 pada residu tiol yang berada di IVR Keap1 mengganggu interaksi dengan Nrf2 menyebabkan misalignment residu lisin dalam Nrf2 yang tidak dapat lagi poliubiquitinylated dan (2) model di mana modifikasi tiol menyebabkan disosiasi Cul3 dari Keap1 [3]. Dalam kedua model, Keap2 yang dimodifikasi oleh induser dan terikat Nrf1 tidak aktif dan, akibatnya, protein Nrf2 yang baru disintesis melewati Keap1 dan mentranslokasi ke dalam nukleus, mengikat ke ARE dan mendorong ekspresi gen target Nrf2 seperti NAD(P)H kuinon oksidoreduktase 1 (NQO1), heme oksigenase 1 (HMOX1), glutamat-sistein ligase (GCL) dan glutathione S transferases (GSTs) (Gbr. 2). Selain modifikasi tiol Keap1 yang menghasilkan induksi gen target Nrf2, protein seperti p21 dan p62 dapat mengikat Nrf2 atau Keap1 sehingga mengganggu interaksi antara Nrf2 dan Keap1 [1], [3] (Gbr. 3).
Gambar 1. Struktur Nrf2 dan Keap1 dan kode sistein. (A) Nrf2 terdiri dari 589 asam amino dan memiliki enam domain yang sangat terkonservasi secara evolusioner, Neh1-6. Neh1 berisi motif bZip, struktur region leucine zipper (L-Zip) dasar, di mana region dasar bertanggung jawab untuk pengenalan DNA dan L-Zip menengahi dimerisasi dengan protein Maf kecil. Neh6 berfungsi sebagai degron untuk memediasi degradasi Nrf2 di dalam nukleus. Neh4 dan 5 adalah domain transaktivasi. Neh2 mengandung motif ETGE dan DLG, yang diperlukan untuk interaksi dengan Keap1, dan daerah hidrofilik residu lisin (7 K), yang sangat diperlukan untuk polubiquitinasi dan degradasi Nrf1 yang bergantung pada Keap2. (B) Keap1 terdiri dari 624 residu asam amino dan memiliki lima domain. Dua motif interaksi protein protein yaitu domain BTB dan domain Kelch dipisahkan oleh intervening region (IVR). Domain BTB bersama dengan bagian N-terminal dari IVR memediasi homodimerisasi Keap1 dan mengikat dengan Cullin3 (Cul3). Domain Kelch dan wilayah C-terminal memediasi interaksi dengan Neh2. (C) Nrf2 berinteraksi dengan dua molekul Keap1 melalui motif Neh2 ETGE dan DLG. Baik ETGE maupun DLG mengikat situs serupa di permukaan bawah motif Keap1 Kelch. (D) Keap1 kaya akan residu sistein, dengan 27 sistein dalam protein manusia. Beberapa sistein ini terletak di dekat residu basa dan oleh karena itu merupakan target elektrofil dan oksidan yang sangat baik. Pola modifikasi residu sistein oleh elektrofil dikenal sebagai kode sistein. Hipotesis kode sistein mengusulkan bahwa agen pengaktif Nrf2 yang berbeda secara struktural mempengaruhi sistein Keap1 yang berbeda. Modifikasi sistein menyebabkan perubahan konformasi di Keap1 mengganggu interaksi antara domain Nrf2 DLG dan Keap1 Kelch, sehingga menghambat polubiquitinasi Nrf2. Pentingnya fungsional Cys151, Cys273 dan Cys288 telah ditunjukkan, karena Cys273 dan Cys288 diperlukan untuk menekan Nrf2 dan Cys151 untuk aktivasi Nrf2 oleh penginduksi [1], [3].
Gambar. 2. Jalur pensinyalan Nrf2-Keap1. (A dan B) dalam kondisi basal, dua molekul Keap1 berikatan dengan Nrf2 dan Nrf2 adalah polyubiquitylated oleh kompleks ligase E3 yang berbasis CulxNUMX. Poliubiquitilasi ini menghasilkan degradasi Nrf3 yang cepat oleh proteasome. Sebagian kecil Nrf2 keluar dari kompleks penghambatan dan terakumulasi dalam nukleus untuk memediasi ekspresi gen tergantung-basis AS, sehingga mempertahankan homeostasis seluler. (C) Dalam kondisi stres, induser memodifikasi cysteines Keap2 yang mengarah ke penghambatan Nrf1 ubiquitylation melalui disosiasi kompleks penghambatan. (D) Menurut model engsel dan latch, modifikasi residu cysteine Keap2 yang spesifik mengarah pada perubahan konformasi dalam Keap1 yang menghasilkan pelepasan motif Nrf1 DLG dari Keap2. Ubiquitination of Nrf1 terganggu tetapi pengikatan dengan motif ETGE tetap ada. (E) Dalam model disosiasi Keap2-Cul1, pengikatan Keap3 dan Cul1 terganggu sebagai respons terhadap elektrofil, yang mengarah ke pelarian Nrf3 dari sistem ubiquitinasi. Dalam kedua model yang disarankan, Keap2 yang diinduksi oleh peredam dan Nrf2 tidak aktif dan, akibatnya, protein Nrf1 yang baru disintesis memotong Keap2 dan mentranslokasi ke dalam nukleus, mengikat ke Antioksidan Respons Element (ARE) dan mendorong ekspresi target Nrf1 gen seperti NQO2, HMOX1, GCL, dan GST [1], [1].
Gambar. 3. Mekanisme untuk akumulasi nuklir konstitutif Nrf2 pada kanker. (A) Mutasi somatik dalam Nrf2 atau Keap1 mengganggu interaksi dua protein ini. Dalam Nrf2, mutasi mempengaruhi motif ETGE dan DLG, tetapi dalam mutasi Keap1 terdistribusi lebih merata. Selanjutnya, aktivasi onkogen, seperti KrasG12D [5], atau gangguan penekan tumor, seperti PTEN [11] dapat menyebabkan induksi transkripsi Nrf2 dan peningkatan Nrf2 nuklir. (B) Hypermethylation dari promotor Keap1 di paru-paru dan kanker prostat mengarah ke pengurangan ekspresi mRNA Keap1, yang meningkatkan akumulasi nuklir Nrf2 [6], [7]. (C) Pada karsinoma ginjal papilaris familial, hilangnya aktivitas enzim fumarat hidratase mengarah pada akumulasi fumarat dan selanjutnya ke suksinat residu cysteine Keap1 (2SC). Modifikasi pasca-translasi ini mengarah ke gangguan interaksi Keap1-Nrf2 dan akumulasi nuklir Nrf2 [8], [9]. (D) Akumulasi protein pengganggu seperti p62 dan p21 dapat mengganggu ikatan Nrf2-Keap1 dan menghasilkan peningkatan Nrf2 nuklir. p62 berikatan dengan Keap1 yang tumpang tindih dengan kantung pengikat untuk Nrf2 dan p21 secara langsung berinteraksi dengan motif DLG dan ETGE dari Nrf2, sehingga bersaing dengan Keap1 [10].
Mekanisme Aktivasi dan Disregulasi Nrf2 di Kanker
Meskipun sitoproteksi yang disediakan oleh aktivasi Nrf2 penting untuk kemoprevensi kanker pada jaringan normal dan premaligna, pada sel yang sepenuhnya ganas, aktivitas Nrf2 memberikan keuntungan pertumbuhan dengan meningkatkan kemoresisten kanker dan meningkatkan pertumbuhan sel tumor [4]. Beberapa mekanisme dimana jalur pensinyalan Nrf2 secara konstitutif diaktifkan dalam berbagai kanker telah dijelaskan: (1) mutasi somatik di Keap1 atau domain pengikatan Keap1 Nrf2 mengganggu interaksi mereka; (2) membungkam epigenetik dari ekspresi Keap1 yang mengarah ke penindasan Nrf2 yang cacat; (3) akumulasi protein pengganggu seperti p62 yang menyebabkan disosiasi kompleks Keap1-Nrf2; (4) induksi transkripsi Nrf2 oleh onkogenik K-Ras, B-Raf dan c-Myc; dan (5) modifikasi pasca-translasi cysteines Keap1 oleh suksinilasi yang terjadi pada karsinoma ginjal papiler familial karena hilangnya aktivitas enzim fumarat hidratase [3], [4], [5], [6], [7], [ 8], [9], [10] (Gbr. 3). Protein Nrf2 yang berlimpah secara meluas menyebabkan meningkatnya ekspresi gen yang terlibat dalam metabolisme obat sehingga meningkatkan resistensi terhadap obat-obat kemoterapi dan radioterapi. Selain itu, tingkat protein Nrf2 tinggi dikaitkan dengan prognosis buruk pada kanker [4]. Nrf2 yang terlalu aktif juga mempengaruhi proliferasi sel dengan mengarahkan glukosa dan glutamin menuju jalur anabolik yang menambah sintesis purin dan mempengaruhi jalur pentosa fosfat untuk mendorong proliferasi sel [11] (Gambar 4).
Gambar. 4. Peran ganda Nrf2 dalam tumorigenesis. Dalam kondisi fisiologis, tingkat Nrf2 nuklir yang rendah cukup untuk pemeliharaan homeostasis seluler. Nrf2 menghambat inisiasi tumor dan metastasis kanker dengan menghilangkan karsinogen, ROS dan agen pengurai DNA lainnya. Selama tumorigenesis, akumulasi kerusakan DNA menyebabkan hiperaktivitas konstitutif Nrf2 yang membantu sel-sel ganas otonom untuk bertahan tingkat tinggi ROS endogen dan untuk menghindari apoptosis. Tingkat Nrf2 nuklir yang terus-menerus meningkat mengaktifkan gen-gen metabolik selain gen-gen sitoprotektor yang berkontribusi pada pemrograman ulang metabolik dan peningkatan proliferasi sel. Kanker dengan tingkat Nrf2 tinggi dikaitkan dengan prognosis yang buruk karena radio dan kemoresistance dan proliferasi sel kanker yang agresif. Dengan demikian, aktivitas jalur Nrf2 bersifat protektif pada tahap awal tumorigenesis, tetapi merugikan pada tahap selanjutnya. Oleh karena itu, untuk pencegahan kanker, meningkatkan aktivitas Nrf2 tetap merupakan pendekatan penting sedangkan untuk pengobatan kanker, penghambatan Nrf2 diinginkan [4], [11].
Mengingat aktivitas Nrf2 yang tinggi umumnya terjadi pada sel kanker dengan hasil yang merugikan, ada kebutuhan untuk terapi untuk menghambat Nrf2. Sayangnya, karena kesamaan struktural dengan beberapa anggota keluarga bZip lainnya, pengembangan inhibitor Nrf2 spesifik adalah tugas yang menantang dan hanya beberapa studi penghambatan Nrf2 yang telah dipublikasikan hingga saat ini. Dengan menyaring produk alami, Ren et al. [12] mengidentifikasi senyawa bradenol antineoplastik sebagai inhibitor Nrf2 yang meningkatkan kemanjuran kemoterapi dari cisplatin. Selain itu, inhibitor PI3K [11], [13] dan Nrf2 siRNA [14] telah digunakan untuk menghambat Nrf2 dalam sel kanker. Baru-baru ini, kami telah menggunakan pendekatan alternatif, yang dikenal sebagai terapi gen kanker bunuh diri, untuk menargetkan sel kanker dengan tingkat Nrf2 tinggi. Nrf2-driven lentiviral vector [15] yang mengandung thymidine kinase (TK) ditransfer ke sel kanker dengan aktivitas ARE tinggi dan sel-sel diobati dengan pro-obat, gansiklovir (GCV). GCV dimetabolisme menjadi GCV-monofosfat, yang selanjutnya terfosforilasi oleh kinase seluler menjadi bentuk trifosfat beracun [16] (Gambar 5). Ini mengarah pada pembunuhan yang efektif tidak hanya TK yang mengandung sel-sel tumor, tetapi juga sel-sel tetangga karena efek pengamat [17]. Terapi gen TK / GCV yang diatur oleh peraturan dapat ditingkatkan lebih lanjut melalui penggabungan agen kemoterapis kanker dengan doxorubicin ke pengobatan [16], yang mendukung gagasan bahwa pendekatan ini dapat berguna dalam hubungannya dengan terapi tradisional.
Gambar. 5. Terapi gen bunuh diri. Penumpukan nuklir Nrf2 pada sel kanker dapat dimanfaatkan dengan menggunakan vektor virus Nrf2 untuk terapi kanker bunuh diri kanker [16]. Dalam pendekatan ini, lentiviral vector (LV) mengekspresikan thymidine kinase (TK) di bawah promotor SV40 minimal dengan empat AREs ditransduksi ke sel adenokarsinoma paru. Tingkat Nrf2 nuklir yang tinggi menyebabkan ekspresi TK yang kuat melalui pengikatan Nrf2. Sel-sel kemudian diobati dengan pro-obat, gansiklovir (GCV), yang difosforilasi oleh TK. Triphosphorylated GCV mengganggu sintesis DNA dan mengarah ke pembunuhan yang efektif tidak hanya TK yang mengandung sel-sel tumor, tetapi juga sel-sel tetangga karena efek pengamat.
Nrf2 adalah pengatur utama yang memicu produksi antioksidan kuat dalam tubuh manusia yang membantu menghilangkan stres oksidatif. Berbagai enzim antioksidan, seperti superoksida dismutase, atau SOD, glutathione, dan katalase, juga diaktifkan melalui jalur Nrf2. Selain itu, phytochemical tertentu seperti kunyit, ashwagandha, bacopa, teh hijau, dan milk thistle, aktifkan Nrf2. Studi penelitian telah menemukan itu Aktivasi Nrf2 secara alami dapat meningkatkan perlindungan seluler dan mengembalikan keseimbangan ke tubuh manusia.
Dr Alex Jimenez DC, CCST Insight
Sulforaphane dan Dampaknya pada Kanker, Kematian, Penuaan, Otak dan Perilaku, Penyakit Jantung & Lainnya
Isothiocyanate adalah beberapa senyawa tanaman yang paling penting yang bisa Anda dapatkan dalam diet Anda. Dalam video ini saya membuat kasus yang paling komprehensif untuk mereka yang pernah dibuat. Rentang perhatian yang pendek? Lewati ke topik favorit Anda dengan mengklik salah satu poin waktu di bawah ini. Garis waktu penuh di bawah ini.
Bagian utama:
00: 01: 14 - Kanker dan kematian
00: 19: 04 - Penuaan
00: 26: 30 - Otak dan perilaku
00: 38: 06 - Rekap terakhir
00: 40: 27 - Dosis
Lini waktu penuh:
00: 00: 34 - Pengantar sulforaphane, fokus utama dari video.
00: 01: 14 - konsumsi sayuran Cruciferous dan pengurangan dalam semua penyebab kematian.
00: 02: 12 - Risiko kanker prostat.
00: 02: 23 - Risiko kanker kandung kemih.
00: 02: 34 - Kanker paru-paru berisiko pada perokok.
00: 02: 48 - Risiko kanker payudara.
00: 03: 13 - Hipotetis: bagaimana jika Anda sudah menderita kanker? (intervensi)
00: 03: 35 - Mekanisme yang masuk akal mengemudi kanker dan data asosiatif kematian.
00: 04: 38 - Sulforaphane dan kanker.
00: 05: 32 - Hewan bukti yang menunjukkan efek kuat dari ekstrak kecambah brokoli pada perkembangan tumor kandung kemih pada tikus.
00: 06: 06 - Pengaruh suplementasi langsung sulforaphane pada pasien kanker prostat.
00: 07: 09 - Bioakumulasi metabolit isothiocyanate dalam jaringan payudara yang sebenarnya.
00: 08: 32 - Penghambatan sel induk kanker payudara.
00: 08: 53 - Pelajaran sejarah: brassica ditetapkan memiliki sifat-sifat kesehatan bahkan di Roma kuno.
00: 09: 16 - Kemampuan Sulforaphane untuk meningkatkan ekskresi karsinogen (benzena, akrolein).
00: 09: 51 - NRF2 sebagai saklar genetik melalui elemen respons antioksidan.
00: 10: 10 - Bagaimana aktivasi NRF2 meningkatkan ekskresi karsinogen melalui glutathione-S-conjugates.
00: 10: 34 - kubis Brussel meningkatkan glutathione-S-transferase dan mengurangi kerusakan DNA.
00: 11: 20 - Broccoli sprout drink meningkatkan ekskresi benzena oleh 61%.
00: 13: 31 - Broccoli sprout homogenate meningkatkan enzim antioksidan di saluran napas bagian atas.
00: 15: 45 - konsumsi sayuran Cruciferous dan kematian penyakit jantung.
00: 16: 55 - Bubuk tunas brokoli meningkatkan lipid darah dan risiko penyakit jantung secara keseluruhan pada penderita diabetes tipe 2.
00: 19: 04 - Awal dari bagian penuaan.
00: 19: 21 - Diet yang diperkaya Sulforaphane meningkatkan masa hidup kumbang dari 15 ke 30% (dalam kondisi tertentu).
00: 20: 34 - Pentingnya peradangan rendah untuk umur panjang.
00: 22: 05 - Sayuran dan tunas kecambah brokoli tampaknya mengurangi beragam penanda inflamasi pada manusia.
00: 23: 40 - Rekap video pertengahan: kanker, bagian penuaan
00: 24: 14 - Studi pada tikus menunjukkan sulforaphane dapat meningkatkan fungsi imun adaptif di usia tua.
00: 25: 18 - Sulforaphane meningkatkan pertumbuhan rambut pada model tikus botak. Gambar di 00: 26: 10.
00: 26: 30 - Awal dari bagian otak dan perilaku.
00: 27: 18 - Pengaruh ekstrak kecambah brokoli pada autisme.
00: 27: 48 - Pengaruh glucoraphanin pada skizofrenia.
00: 28: 17 - Mulai dari diskusi depresi (mekanisme dan studi yang masuk akal).
00: 31: 21 - Studi mouse menggunakan 10 model yang berbeda dari depresi yang diinduksi stres menunjukkan sulforaphane sama efektifnya dengan fluoxetine (prozac).
00: 32: 00 - Studi menunjukkan konsumsi langsung glukoraphanin pada tikus juga efektif mencegah depresi dari model stres kekalahan sosial.
00: 33: 01 - Awal dari bagian neurodegenerasi.
00: 33: 30 - Sulforaphane dan penyakit Alzheimer.
00: 33: 44 - Sulforaphane dan penyakit Parkinson.
00: 33: 51 - Sulforaphane dan penyakit Hungtington.
00: 34: 13 - Sulforaphane meningkatkan protein heat shock.
00: 34: 43 - Awal dari bagian cedera otak traumatis.
00: 35: 01 - Sulforaphane disuntikkan segera setelah TBI meningkatkan daya ingat (studi pada tikus).
00: 35: 55 - Sulforaphane dan plastisitas neuronal.
00: 36: 32 - Sulforaphane meningkatkan pembelajaran dalam model diabetes tipe II pada tikus.
00: 37: 19 - Sulforaphane dan duchenne muscular dystrophy.
00: 37: 44 - Myostatin inhibition dalam sel-sel satelit otot (in vitro).
00: 38: 06 - Rekap video-ulang: mortalitas dan kanker, kerusakan DNA, stres oksidatif dan peradangan, ekskresi benzena, penyakit kardiovaskular, diabetes tipe II, efek pada otak (depresi, autisme, skizofrenia, neurodegenerasi), jalur NRF2.
00: 40: 27 - Pikiran tentang mencari tahu kecambah brokoli atau sulforaphane.
00: 41: 01 - Anekdot saat bertumbuh di rumah.
00: 43: 14 - Pada suhu memasak dan aktivitas sulforaphane.
00: 43: 45 - Konversi bakteri usus dari sulforaphane dari glucoraphanin.
00: 44: 24 - Suplemen bekerja lebih baik ketika dikombinasikan dengan myrosinase aktif dari sayuran.
00: 44: 56 - Teknik memasak dan sayuran silangan.
00: 46: 06 - Isothiocyanate sebagai goitrogens.
Ucapan Terima Kasih
Karya ini didukung oleh Akademi Finlandia, Yayasan Sigrid Juselius, dan Organisasi Kanker Finlandia.
Kesimpulannya, faktor nuklir (berasal dari eritroid 2) seperti 2, juga dikenal sebagai NFE2L2 atau Nrf2, adalah protein yang meningkatkan produksi antioksidan yang melindungi tubuh manusia dari stres oksidatif. Seperti dijelaskan di atas, stimulasi jalur Nrf2 sedang dipelajari untuk pengobatan penyakit yang disebabkan oleh stres oksidatif, termasuk kanker. Cakupan informasi kami terbatas pada masalah chiropraktik dan kesehatan tulang belakang. Untuk mendiskusikan materi pelajaran, jangan ragu untuk bertanya kepada Dr. Jimenez atau hubungi kami di�915-850-0900 .
Diskusi Topik Tambahan: Menghilangkan Nyeri Lutut tanpa Pembedahan
Nyeri lutut adalah gejala umum yang dapat terjadi karena berbagai cedera dan / atau kondisi lutut, termasuk cedera olahraga. Lutut adalah salah satu sendi paling kompleks di tubuh manusia karena terdiri dari perpotongan empat tulang, empat ligamen, berbagai tendon, dua menisci, dan tulang rawan. Menurut American Academy of Family Physicians, penyebab paling umum dari nyeri lutut termasuk subluksasi patella, tendinitis patella atau lutut jumper, dan penyakit Osgood-Schlatter. Meskipun nyeri lutut paling mungkin terjadi pada orang di atas usia 60 tahun, nyeri lutut juga dapat terjadi pada anak-anak dan remaja. Nyeri lutut dapat dirawat di rumah mengikuti metode RICE, namun, cedera lutut yang parah mungkin memerlukan perhatian medis segera, termasuk perawatan chiropractic.
DNA mendukung sekitar gen 20,000, masing-masing memegang program untuk penciptaan protein atau enzim yang diperlukan untuk gaya hidup sehat. Setiap pola ini perlu secara konstan diatur oleh semacam "promotor" yang mengelola dengan tepat berapa banyak masing-masing zat dan / atau bahan kimia dihasilkan dan dalam kondisi apa ini juga akan berkembang.
Dengan menghubungkan ke jenis tertentu bidang promotor seperti saklar, yang dikenal sebagai Elemen Respons Antioksidan, atau ARE, Faktor Nrf2Mendukung kecepatan penciptaan ratusan gen berbeda yang memungkinkan sel bertahan dalam keadaan yang penuh tekanan. Gen ini kemudian menghasilkan pilihan enzim antioksidan yang mengembangkan jaringan pertahanan dengan menetralkan oksidan dan dengan membersihkan produk sampingan beracun yang tertinggal dalam produksinya, selain membantu memulihkan kerusakan yang ditimbulkannya.
Apa itu Stres Oksidatif?
Beberapa oksidan seperti radikal superoksida, atau O2-., Dan hidrogen peroksida, atau H2O2, telah dibuat melalui praktik pembakaran zat dan / atau bahan kimia yang menopang tubuh manusia. Tubuh manusia memiliki enzim antioksidan yang menetralkan dan mendetoksifikasi makanan dan minuman reaktif yang kita konsumsi. Nrf2 memodulasi produksinya untuk menjaga keseimbangan dan menggarisbawahi permintaan untuk semua enzim ini. Keseimbangan ini dapat terganggu oleh beberapa faktor, termasuk usia.
Seiring bertambahnya usia, tubuh manusia menciptakan lebih sedikit Nrf2 dan keseimbangan yang halus ini secara bertahap dapat mulai beralih ke sisi oksidatif, keadaan yang disebut sebagai stres oksidatif. Penyakit juga dapat menyebabkan produksi oksidan yang berlebihan. Infeksi, alergi, dan gangguan autoimun juga dapat memicu sel kekebalan kita untuk membuat oksidan reaktif, seperti O2-. , H2O2, OH dan HOCl, dimana sel-sel sehat menjadi rusak dan merespon dengan inflamasi. Penyakit yang terkait dengan penuaan, termasuk serangan jantung, stroke, kanker, dan kondisi neurodegeneratif seperti penyakit Alzheimer, juga meningkatkan perkembangan oksidan, menghasilkan stres dan respons peradangan.
Apa itu Aktivator Nrf2?
Protein Nrf2, juga disebut faktor transkripsi karena cara itu dapat mendukung dan mengontrol enzim dan gen, adalah elemen rahasia dari urutan reaksi biokimia dalam sel yang bereaksi terhadap modifikasi dalam keseimbangan kognitif serta keseimbangan oksidatif. Unsur-unsur penginderaan jalur ini memodifikasi dan mengeluarkan Nrf2, memicunya sehingga mungkin menyebar ke inti sel menuju DNA. Nrf2 dapat mengaktifkan atau mematikan gen dan enzim yang didukungnya untuk melindungi sel.
Untungnya, berbagai zat yang merupakan penggerak Nrf2 berkembang melalui konsumsi tanaman dan ekstrak tertentu yang digunakan berabad-abad yang lalu dalam pengobatan tradisional Cina dan Native American. Phytochemical ini tampaknya sama kuatnya dengan lebih sedikit efek samping, seperti produk farmasi pengaktif Nrf2 yang digunakan saat ini.
Faktor nuklir erythroid 2-faktor terkait, lebih dikenal sebagai Nrf2, adalah faktor transkripsi yang melindungi sel dengan mengatur gen, enzim dan tanggapan antioksidan. Faktor transkripsi adalah sejenis protein yang menempel pada DNA untuk mempromosikan penciptaan zat dan bahan kimia tertentu, termasuk glutation S-transferase, atau GST. Aktivasi Nrf2 menginduksi produksi protein aktif yang menunjukkan kapasitas antioksidan kuat untuk membantu mengurangi stres oksidatif.
Dr Alex Jimenez DC, CCST Insight
Ilmu Di Balik Aktivasi Nrf2
Setelah suplemen makanan pengaktifasi Nrf2 awal dibuat di 2004, informasi minimal diketahui tentang fungsi jalur Nrf2. Sekitar surat kabar 200 dalam literatur tentang Nrf2, juga dikenal sebagai 2 atau NFE2L2, seperti faktor-nuklir, ada dan para peneliti baru saja mulai menemukan respons antioksidan Nrf2 pada mamalia. Pada 2017, bagaimanapun, lebih dari 9,300 studi penelitian akademik tentang "regulator utama," telah dicetak.
Pada kenyataannya, Nrf2 mengatur banyak enzim antioksidan yang tidak berkorelasi dengan gen, sebaliknya, mereka menawarkan perlindungan terhadap berbagai keadaan terkait stres yang ditemui oleh sel, organ dan akhirnya organisme, dalam kondisi sehat dan patologis. Berdasarkan kuantitas informasi baru ini dari studi penelitian akademik yang diterbitkan, para peneliti sekarang dapat berkembang lebih baik Suplemen diet Nrf2.
Pada tahun 2007, studi penelitian telah menunjukkan fungsi kompleks jalur Nrf2. Aktivator Nrf2 ditemukan meniru faktor struktur yang berbeda di dalam tubuh manusia. Melalui jalur ini, aktivator Nrf2 telah diperlengkapi untuk merasakan perubahan kondisi di seluruh sel untuk menjaga keseimbangan dan merespons kebutuhan gen yang berkembang.
Mengapa Menggunakan Suplemen Aktivasi Nrf2?
Karena kemampuan aktivasi Nrf2 berkurang dengan bertambahnya usia dalam organisme, perubahan mungkin mulai terjadi. Studi penelitian telah menunjukkan bahwa fokus Nrf2 dalam sel menurun dengan bertambahnya usia, menunjukkan peningkatan tanda stres oksidatif. Berbagai penyakit terkait usia seperti aterosklerosis dan penyakit kardiovaskular, radang sendi, kanker, obesitas, diabetes tipe 2, hipertensi, katarak, dan penyakit Alzheimer serta penyakit Parkinson dapat berkembang karena perubahan-perubahan ini. Stres oksidatif telah ditemukan pada masalah kesehatan ini.
Dengan merangsang kapasitas sel untuk meningkatkan produksi aktivator Nrf2, Suplemen diet Nrf2 dapat membantu menghidupkan kembali kemampuan tubuh manusia untuk melawan efek dari stres oksidatif. Asam lemak tak jenuh ganda, atau PUFA, adalah salah satu molekul yang paling mudah teroksidasi dan sangat rentan mengalami kerusakan akibat radikal bebas. Asam tiobarbiturat, atau TBARS, produksi dapat meningkat seiring bertambahnya usia, menunjukkan peningkatan stres oksidatif bersama dengan penurunan jalur yang diatur Nrf2.
Secara biologis, induksi gen adalah mekanisme yang sangat lambat, umumnya membutuhkan waktu berjam-jam untuk ditransfer melalui jalur. Akibatnya, banyak enzim memiliki sakelar hidup / mati sendiri yang dapat dipicu dalam hitungan menit oleh enzim pengatur yang berbeda. Para peneliti telah mengembangkan komposisi kepemilikan dari aktivator Nrf2 yang memanfaatkan basis pengetahuan aktivasi ini. Aktivasi Nrf2 tidak hanya terdiri dari faktor transkripsi Nrf2 yang dilepaskan dari inhibitornya dan bermigrasi ke inti sel, tetapi juga mengikat urutan DNA tertentu untuk mendorong ekspresi gen sitoprotektif, mengatur kecepatan saat Nrf2 dikeluarkan dari nukleus.
Memahami prosedur eliminasi dan aktivasi Nrf2 dalam tubuh manusia telah memungkinkan para peneliti untuk membangun kombinasi dari aktivator Nrf2 yang berbeda untuk mencapai refleksi gen melalui modulasinya. Kombinasi dari basis pengetahuan, bersama dengan berbagai studi penelitian lainnya telah membantu menghasilkan aktivator Nrf2 untuk digunakan sebagai suplemen makanan. Cakupan informasi kami terbatas pada masalah chiropraktik dan kesehatan tulang belakang. Untuk membahas pokok bahasan ini, jangan ragu untuk bertanya kepada Dr. Jimenez atau hubungi kami di 915-850-0900 .
Diundangkan oleh Dr. Alex Jimenez
Diskusi Topik Tambahan: Menghilangkan Nyeri Lutut tanpa Pembedahan
Nyeri lutut adalah gejala umum yang dapat terjadi karena berbagai cedera dan / atau kondisi lutut, termasuk cedera olahraga. Lutut adalah salah satu sendi paling kompleks di tubuh manusia karena terdiri dari perpotongan empat tulang, empat ligamen, berbagai tendon, dua menisci, dan tulang rawan. Menurut American Academy of Family Physicians, penyebab paling umum dari nyeri lutut termasuk subluksasi patella, tendinitis patella atau lutut jumper, dan penyakit Osgood-Schlatter. Meskipun nyeri lutut paling mungkin terjadi pada orang di atas usia 60 tahun, nyeri lutut juga dapat terjadi pada anak-anak dan remaja. Nyeri lutut dapat dirawat di rumah mengikuti metode RICE, namun, cedera lutut yang parah mungkin memerlukan perhatian medis segera, termasuk perawatan chiropractic.
Antioksidan secara ilmiah disebut sebagai senyawa yang membatasi proses oksidasi dalam tubuh manusia, yang jika dibiarkan, dapat menciptakan radikal bebas yang dapat mengembangkan banyak reaksi berantai yang dapat menyebabkan kerusakan sel. Untungnya, tubuh manusia dapat menciptakan mekanisme kekebalan bawaan, namun, ketika memasang spesies oksigen reaktif, atau ROS, tidak dapat dinetralkan, membayangkan api kecil yang keluar dari kontrol ketika diinfuskan dengan oksigen, bahaya pasti akan terjadi. .
Untuk terus memperluas pada metafora nyala api, produk akhir dari tidak memiliki kemampuan untuk menetralkan dampak ROS, atau spesies oksigen reaktif, adalah kerusakan serta peradangan, dengan kata lain, tubuh manusia secara harfiah terbakar. Yang luar biasa adalah ada antioksidan yang sangat membantu melawan masalah kesehatan ini dan antioksidan ini adalah glutathione. Meskipun ditemukan di 1889, efek antioksidan glutathione telah menjadi salah satu topik yang paling menarik dalam penelitian modern.
Master Antioksidan: Glutathione
Zat yang kuat adalah tripeptida yang berkembang dari sistein, asam glutamat, dan glisin. Karena kemampuannya untuk melindungi tubuh manusia dari pembentukan radikal bebas, glutathione pada akhirnya dapat membantu meningkatkan sistem kekebalan yang sehat. Berdasarkan Laporan Ilmiah di 2015, ditentukan bahwa kapasitas glutathione untuk berfungsi secara sinergis dengan peroxiredin dan katalase membantu menjaga sel terhadap hidrogen peroksida. Formula sinergis ini berfungsi terhadap spesies oksigen reaktif, atau ROS. Glutathione, peroxidredin dan katalase adalah elemen penting dalam peningkatan homeostasis seluler, yang merupakan proses penting dari sel-sel sehat, jaringan dan organ sama sekali.
Selain itu, glutathione meningkatkan keseluruhan struktur dan fungsi sistem kekebalan tubuh dengan memanfaatkan efeknya yang penting pada fungsi limfosit. Menurut Departemen Imunokimia, menambah kadar glutathione dalam tubuh manusia dengan benar dapat sangat meningkatkan reaksi kekebalan tubuh. Sebagai contoh, dua uji coba terkontrol plasebo acak menunjukkan bahwa pengobatan terapeutik pasien dengan gangguan kekebalan dengan N-asetil-sistein, atau NAC, menghasilkan, pada kedua kasus, dalam pertumbuhan substansial dalam sebagian besar proses imunologi yang termasuk peremajaan keseluruhan. aktivitas sel pembunuh alami. N-acetyl-cysteine, atau NAC, menggunakan sulfur dari glutathione dan menggabungkannya dengan molekul beracun, yang kemudian menjadi larut dalam air dan dibuang di tubuh manusia.
Glutathione juga memiliki kemampuan untuk merevitalisasi lipoic acid serta mendaur ulang Vitamin C dan E, yang diperlukan untuk memulai proses sistem tertentu dengan mengirim elektron untuk menetralisir radikal bebas. Berdasarkan studi penelitian dari PLoS ONE, glutathione mempengaruhi pasien dengan diabetes metilus, atau T2DM, dan mycobacterium tuberculosis. Biasanya, individu dengan sistem kekebalan yang lemah memiliki kecenderungan untuk menunjukkan paparan yang lebih besar terhadap M. tb, atau mycobacterium tuberculosis, penyakit atau infeksi. Lebih lanjut, individu dengan diabetes mellitus tipe 2, atau T2DM, dua sampai tiga kali lebih rentan terhadap TB daripada orang tanpa T2DM. Studi penelitian juga menyarankan bahwa meningkatkan kadar glutathione pada makrofag yang diisolasi dari pasien dengan T2DM menyebabkan peningkatan kontrol penyakit M.Tb atau infeksi. Hasil ini menunjukkan bahwa tingkat glutathione yang lebih rendah pada pasien dengan T2DM berkontribusi pada peningkatan kemungkinan penyakit M. tb atau infeksi. Apalagi tergantung pada Dietro Ghezzi di Brighton dan Sussex Medical School, stres oksidatif akhirnya dapat menyebabkan struktur dan fungsi sistem kekebalan tubuh yang buruk.
Untungnya, glutathione memainkan peran penting dalam memperkuat dan mengendalikan kekebalan. Sebagai contoh, glutathione sangat penting untuk proses bawaan dan adaptif dalam sistem kekebalan, termasuk proliferasi T-limfosit, aktivitas fagositik dari neutrofil polimorfonuklear, dan fungsi sel dendritik, yang dapat menjadi fundamental karena ini terdiri dari sel-sel yang menyajikan antigen. . Imunitas sel-meditated termasuk antigen protein yang awalnya mulai berdegenerasi di vesikula endositik makrofag dan sel dendritik, oleh karena itu, peptida yang lebih kecil ditunjukkan pada permukaan untuk mengaktifkan proliferasi sel T antigen-spesifik. Selain itu, glutathione membantu terciptanya sitokin, dan perlu untuk mempertahankan produksi interferon-gamma oleh sel dendritik, yang penting untuk melindungi terhadap patogen intraseluler termasuk mycobacteria.
N-acetyl-cysteine, atau NAC, secara ilmiah disebut sebagai prekursor glutathione, juga merupakan antioksidan seluler yang sangat kuat digunakan sebagai antioksidan penangkal radikal bebas. Biasa dikenal karena perannya dalam menghindari toksisitas acetaminophen, NAC, atau N-acetyl-cysteine, telah dibuktikan memiliki beberapa manfaat kesehatan dan kebugaran. Berdasarkan Jurnal Sel, NAC membantu mendukung respons peradangan yang sehat dan dapat berdampak positif pada tenaga kerja jangka panjang dan prematur. Studi penelitian menyimpulkan bahwa pada wanita dengan kelahiran prematur sebelumnya dan vaginosis bakterial, 0.6 gram NAC per hari diambil secara oral bersama dengan progesteron setelah minggu 16 kehamilan terlindung terhadap kekambuhan kelahiran prematur dan meningkatkan hasil neonatal. Kesimpulannya, efek positif NAC pada pembentukan otot juga terdeteksi. Setelah tiga menit kontraksi persisten, ada peningkatan output 15 persen, menunjukkan bagaimana NAC memainkan peran mendasar dalam meningkatkan pembentukan otot dan mengurangi kelelahan secara keseluruhan selama persalinan.
Para peneliti juga menemukan bahwa NAC, atau N-acetyl-cysteine, dapat bermanfaat bagi mereka yang memiliki sindrom ovarium polikistik, atau PCOS. PCOS, atau sindrom ovarium polikistik, adalah penyakit terkait kelenjar endokrin yang umum terjadi yang memengaruhi sekitar 5 hingga 10 persen wanita usia reproduksi. Pada pasien seperti itu, ada risiko lebih besar untuk mengalami sindrom metabolik, dimana penggunaan NAC membantu memulihkan kadar dan sensitivitas insulin yang sehat.
Wawasan Dr. Alex Jimenez
Glutathione telah disebut sebagai "master antioksidan" karena peran fundamentalnya dalam mencapai dan menjaga kesehatan dan kebugaran secara keseluruhan. Sementara tubuh manusia mampu menghasilkan glutathione sendiri, nutrisi yang buruk, polusi, racun, penggunaan obat-obatan dan / atau obat-obatan yang berlebihan, stres, trauma, penuaan, penyakit dan radiasi dapat menurunkan tingkat glutathione alami kita. Hal ini pada gilirannya dapat membuat individu lebih rentan terhadap kerusakan sel dari stres oksidatif, radikal bebas, infeksi dan kanker. Suplementasi Glutathione dapat memiliki manfaat yang luar biasa pada tubuh manusia. Bersama dengan opsi perawatan alternatif, seperti perawatan chiropractic, kadar glutathione sekali lagi dapat diatur untuk meningkatkan kesejahteraan.
Selain itu, para profesional perawatan kesehatan telah menyarankan untuk menerapkan penggunaan suplementasi glutathione bersama dengan pilihan perawatan alternatif lainnya, seperti perawatan chiropractic, untuk lebih meningkatkan kesehatan dan kebugaran secara keseluruhan. Antioksidan penting untuk menjaga kesehatan maksimal serta menghambat reaksi berantai radikal bebas yang menyebabkan kerusakan atau kerusakan sel. Antioksidan kuat seperti glutathione, seperti yang disebutkan di atas, pada akhirnya membantu mengatur perkembangan radikal bebas ini dan memberikan respons sistem kekebalan tubuh yang lebih sehat. Studi penelitian telah menemukan itu perawatan chiropractic mungkin juga memainkan peran penting dalam proses ini, secara alami meningkatkan aktivitas antioksidan dalam tubuh manusia. Perawatan Chiropractic adalah pendekatan pengobatan yang aman dan efektif yang menggunakan penyesuaian tulang belakang dan manipulasi manual untuk memperbaiki misalignment tulang belakang, atau subluksasi, untuk memungkinkan tubuh manusia untuk menyembuhkan dirinya secara alami tanpa menggunakan obat / obat dan / atau intervensi bedah.
Akhirnya, antioksidan menunjukkan sifat biologis mereka melalui banyak manfaat kesehatan, yang mungkin diperlukan sekarang lebih dari sebelumnya dengan semakin meningkatnya serangan stres, penyakit, dan polusi di dunia modern kita, yang semuanya berkontribusi pada kerusakan dan / atau kerusakan sel. . Glutathione dan prekursornya, NAC, atau N-acetyl-cysteine, terus menunjukkan statusnya yang kuat di bidang antioksidan. Bersama dengan pilihan pengobatan alternatif, seperti perawatan kiropraktik, orang dapat memanfaatkan semua manfaat yang ditawarkan antioksidan kuat ini. Cakupan informasi kami terbatas pada chiropraktik serta cedera dan kondisi tulang belakang. Untuk membahas pokok bahasan ini, jangan ragu untuk bertanya kepada Dr. Jimenez atau hubungi kami di 915-850-0900 .
Diundangkan oleh Dr. Alex Jimenez
Topik Tambahan: Back Pain
Nyeri punggung adalah salah satu penyebab utama kecacatan dan hari-hari yang terlewatkan di dunia kerja. Nyatanya, nyeri punggung telah dianggap sebagai alasan paling umum kedua untuk kunjungan ke dokter, hanya kalah jumlah oleh infeksi saluran pernapasan atas. Sekitar 80 persen populasi akan mengalami beberapa jenis nyeri punggung setidaknya sekali sepanjang hidup mereka. Tulang belakang adalah struktur kompleks yang terdiri dari tulang, sendi, ligamen dan otot, di antara jaringan lunak lainnya. Karena ini, cedera dan / atau kondisi yang diperburuk, seperti cakram hernia, akhirnya dapat menyebabkan gejala nyeri punggung. Cedera olahraga atau cedera kecelakaan mobil sering menjadi penyebab paling sering dari nyeri punggung, namun terkadang gerakan yang paling sederhana dapat memiliki hasil yang menyakitkan. Untungnya, pilihan pengobatan alternatif, seperti perawatan chiropractic, dapat membantu meringankan nyeri punggung melalui penggunaan penyesuaian tulang belakang dan manipulasi manual, yang pada akhirnya meningkatkan pereda nyeri.
Ilmu pengetahuan berbasis Chiropractor Dr. Alexander Jimenez lihatlah stres oksidatif, apa itu, bagaimana hal itu mempengaruhi tubuh dan antioksidan pertahanan untuk memperbaiki situasi.
Esra Birben PhD, 1 Umit Murat Sahiner MD, 1 Cansin Sackesen MD, 1 Serpil Erzurum MD, 2 dan Omer Kalayci, MD1
Abstrak: Spesies oksigen reaktif (ROS) diproduksi oleh organisme hidup sebagai hasil metabolisme sel normal dan faktor lingkungan, seperti polutan udara atau asap rokok. ROS adalah molekul yang sangat reaktif dan dapat merusak struktur sel seperti karbohidrat, asam nukleat, lipid, dan protein serta mengubah fungsinya. Pergeseran keseimbangan antara oksidan dan antioksidan yang mendukung oksidan disebut 'stres oksidatif'. Regulasi reduksi dan oksidasi (redoks) sangat penting untuk kelangsungan hidup sel, aktivasi, proliferasi, dan fungsi organ. Organisme aerobik memiliki sistem antioksidan terintegrasi, yang mencakup antioksidan enzimatik dan non-enzimatik yang biasanya efektif dalam memblokir efek berbahaya dari ROS. Namun, dalam kondisi patologis, sistem antioksidan bisa kewalahan. Stres oksidatif berkontribusi pada banyak kondisi dan penyakit patologis, termasuk kanker, gangguan neurologis, aterosklerosis, hipertensi, iskemia / perfusi, diabetes, sindrom gangguan pernapasan akut, fibrosis paru idiopatik, penyakit paru obstruktif kronik, dan asma. Dalam ulasan ini, kami merangkum oksidan seluler dan sistem antioksidan dan membahas efek seluler dan mekanisme stres oksidatif.
Kata kunci: antioksidan, oksidan, stres oksidatif, spesies oksigen reaktif, redoks
(Jurnal WAO 2012; 5: 9 19)
Spesies oksigen reaktif (ROS) diproduksi oleh organisme hidup sebagai hasil dari metabolisme sel normal. Pada konsentrasi rendah hingga sedang, mereka berfungsi dalam proses fisiologis sel, tetapi pada konsentrasi tinggi, mereka menghasilkan modifikasi yang merugikan pada komponen sel, seperti lipid, protein, dan DNA.1�6 Pergeseran keseimbangan antara oksidan/antioksidan yang mendukung oksidan disebut "stres oksidatif." Stres oksidatif berkontribusi pada banyak kondisi patologis, termasuk kanker, gangguan neurologis,7-10 aterosklerosis, hipertensi, iskemia/perfusi,11-14 diabetes, sindrom gangguan pernapasan akut, fibrosis paru idiopatik, penyakit paru obstruktif kronik ,15 dan asma.16-21 Organisme aerobik memiliki sistem antioksidan terintegrasi,� yang meliputi antioksidan enzimatik dan nonenzimatik yang biasanya efektif dalam memblokir efek berbahaya dari ROS. Namun, dalam kondisi patologis, sistem antioksidan dapat kewalahan. Dalam ulasan ini, kami merangkum sistem oksidan dan antioksidan seluler dan regulasi keadaan reduksi dan oksidasi (redoks) dalam keadaan sehat dan sakit.
OXIDANTS
Sumber Endogen dari ROS
ROS diproduksi dari molekul oksigen sebagai hasil metabolisme sel normal. ROS dapat dibagi menjadi kelompok 2: radikal bebas dan nonradis. Molekul yang mengandung satu atau lebih elektron tak berpasangan dan dengan demikian memberi reaktivitas terhadap molekul disebut radikal bebas. Bila radikal bebas 2 membagikan elektron mereka yang tidak berpasangan, bentuk non-statis telah dibuat. ROS utama 3 yang bermakna fisiologis adalah anion superoksida (O22.), Radikal hidroksil (OH), dan hydro-gen peroxide (H2O2). ROS dirangkum dalam Tabel 1.
Anion superoksida dibentuk dengan penambahan 1 elektron ke molekul oksigen.22 Proses ini dimediasi oleh nikotin adenin dinukleotida fosfat [NAD(P)H] oksidase atau xantin oksidase atau oleh sistem transpor elektron mitokondria. Tempat utama untuk memproduksi anion superoksida adalah mitokondria, mesin sel untuk memproduksi adenosin trifosfat. Biasanya, elektron ditransfer melalui rantai transpor elektron mitokondria untuk mereduksi oksigen menjadi air, tetapi sekitar 1 hingga 3% dari semua elektron bocor dari sistem dan menghasilkan superoksida. NAD(P)H oksidase ditemukan pada leukosit polimorfonuklear, monosit, dan makrofag. Setelah fagositosis, sel-sel ini menghasilkan ledakan superoksida yang menyebabkan aktivitas bakterisida. Superoksida diubah menjadi hidrogen peroksida oleh aksi dismutase superoksida (SOD, EC 1.15.1.1). Hidrogen peroksida mudah berdifusi melintasi membran plasma. Hidrogen peroksida juga diproduksi oleh xantin oksidase, asam amino oksidase, dan NAD(P)H oksidase�23,24 dan dalam peroksisom dengan konsumsi oksigen molekuler dalam reaksi metabolisme. Dalam serangkaian reaksi yang disebut reaksi Haber�Weiss dan Fenton, H2O2 dapat terurai menjadi OH2 dengan adanya logam transmisi seperti Fe21 atau Cu21.25
Fe31 +�.O2�?Fe2 +�O2 Haber Weiss
Fe2 +�H2O2�?Fe3 +�OH�+ .OH Reaksi Fenton
O 2 sendiri juga dapat bereaksi dengan H2 O2 dan menghasilkan OH.26,27 Radikal hidroksil merupakan ROS yang paling reaktif dan dapat merusak protein, lipid, serta karbohidrat dan DNA. Itu juga dapat memulai peroksidasi lipid dengan mengambil elektron dari asam lemak tak jenuh ganda.
Enzim granulositik memperluas reaktivitas H2O2 melalui eosinofil peroksidase dan myeloperoksidase (MPO). Dalam neutrofil aktif, H2O2 dikonsumsi oleh MPO. Dengan adanya ion klorida, H2O2 diubah menjadi asam hipoklorit (HOCl). HOCl sangat oksidatif dan memainkan peran penting dalam membunuh patogen di saluran udara.28 Namun, HOCl juga dapat bereaksi dengan DNA dan menginduksi interaksi protein-DNA dan menghasilkan produk oksidasi pirimidin dan menambahkan klorida ke basa DNA.29,30 Eosinofil peroksidase dan MPO juga berkontribusi pada stres oksidatif dengan modifikasi protein melalui halogenasi, nitrasi, dan ikatan silang protein melalui radikal tirosil.31 33
Radikal radikal lain yang berasal dari oksigen adalah radikal peroksil (ROO $). Bentuk paling sederhana dari radikal ini adalah radikal hidro- peroksil (HOO $) dan memiliki peran dalam peroksidasi asam lemak. Radikal bebas dapat memicu reaksi rantai peroksidasi lipid dengan mengabstraksikan atom hidrogen dari karbon metilen rantai samping. Radikal lipida kemudian bereaksi dengan oksigen untuk menghasilkan radikal peroksil. Radikal peroksil memulai reaksi berantai dan mengubah asam lemak tak jenuh ganda menjadi hydroperoxides lipid. Hydroperoxides lipida sangat tidak stabil dan mudah terurai menjadi produk sekunder, seperti aldehida (seperti 4-hydroxy-2,3-nonenal) dan malondialdehydes (MDA). Isoprostan adalah kelompok produk peroksidasi lipid lain yang dihasilkan melalui peroksidasi asam arakidonat dan juga ditemukan meningkat pada kondensat plasma dan nafas penderita asma.34,35 Peroksidasi lipid mengganggu integritas membran sel dan menyebabkan penataan kembali struktur membran. .
Hidrogen peroksida, radikal superoksida, glutathione teroksidasi (GSSG), MDA, isoprostan, karbonil, dan nitrotyrosin dapat dengan mudah diukur dari sampel lavage plasma, darah, atau bronchoalveolar sebagai biomarker oksidasi dengan uji standar.
Sumber Eksim Oksidan
Asap rokok
Asap rokok mengandung banyak oksidan dan radikal bebas dan senyawa organik, seperti superoksida dan oksida nitrat. 36 Selain itu, menghirup asap rokok ke paru-paru juga mengaktifkan beberapa mekanisme endogen, seperti akumulasi neutrofil dan makrofag, yang selanjutnya meningkatkan cedera oksidan. .
Paparan Ozon
Paparan ozon dapat menyebabkan peroksidasi lipid dan menyebabkan masuknya neutrofil ke epitel saluran napas. Paparan jangka pendek terhadap ozon juga menyebabkan pelepasan mediator inflamasi, seperti MPO, protein kationik eosinofilik dan juga dehidrogenase laktat dan albumin.37 Bahkan pada subyek sehat, paparan ozon menyebabkan pengurangan fungsi paru.38 Cho et al39 telah menunjukkan bahwa partikulat (campuran partikel padat dan tetesan cairan yang tersuspensi di udara) mengkatalisis pengurangan oksigen.
Hyperoxia
Hyperoxia mengacu pada kondisi tingkat oksigen yang lebih tinggi daripada tekanan parsial oksigen normal di paru-paru atau jaringan tubuh lainnya. Ini menyebabkan produksi yang lebih besar dari spesies oksigen dan nitrogen reaktif. 40,41
Radiasi pengion
Radiasi pengion, dengan adanya O2, mengubah radikal hidroksil, superoksida, dan radikal organik menjadi hidrogen peroksida dan hidroperoksida organik. Spesies hidroperoksida ini bereaksi dengan ion logam aktif redoks, seperti Fe dan Cu, melalui reaksi Fenton dan dengan demikian menyebabkan stres oksidatif.42,43 Narayanan et al44 menunjukkan bahwa fibroblas yang terpapar partikel alfa mengalami peningkatan yang signifikan pada O2 2. dan H2O2 intraseluler. produksi melalui membran plasma NADPH oksidase.44 Molekul transduksi sinyal, seperti sinyal ekstraseluler-diatur kinase 1 dan 2 (ERK1 / 2), c-Jun N-terminal kinase (JNK), dan p38, dan faktor transkripsi, seperti aktivator protein-1 (AP-1), faktor nuklir-kB (NF-kB), dan p53, diaktifkan, yang menghasilkan ekspresi gen yang terkait dengan respons radiasi. 45 50 Foton ultraviolet A (UVA) memicu reaksi oksidatif dengan eksitasi fotosensitizer endogen, seperti porfirin, NADPH oksidase, dan riboflavin. 8-Oxo-7,8- dihydroguanine (8-oxoGua) adalah produk oksidasi DNA bermediasi UVA utama yang dibentuk oleh oksidasi radikal OH, oksidan 1-elektron, dan oksigen singlet yang terutama bereaksi dengan guanin.51 Pembentukan guanin kation radikal dalam DNA yang terisolasi telah terbukti terjadi secara efisien melalui efek langsung radiasi pengion.52,53 Setelah terpapar radiasi pengion, kadar glutathione (GSH) intraseluler menurun untuk jangka pendek tetapi kemudian meningkat lagi.54
Ion Logam Berat
Ion logam berat, seperti besi, tembaga, kadmium, merkuri, nikel, timbal, dan arsenik, dapat menyebabkan pembentukan radikal reaktif dan menyebabkan kerusakan sel akibat penipisan aktivitas enzim melalui peroksidasi dan reaksi lipid dengan protein nuklir dan DNA.55
Salah satu mekanisme terpenting pembentukan radikal bebas bermediasi logam adalah melalui reaksi tipe Fenton. Ion superoksida dan hidrogen peroksida dapat berinteraksi dengan logam transisi, seperti besi dan tembaga, melalui reaksi katalis logam Haber Weiss / Fenton membentuk radikal OH.
Selain mekanisme tipe Fenton dan Haber-Weiss, ion logam tertentu dapat bereaksi langsung dengan molekul seluler untuk menghasilkan radikal bebas, seperti radikal tiol, atau menginduksi jalur pensinyalan sel. Radikal ini juga dapat bereaksi dengan molekul tiol lain untuk menghasilkan O22 .. O22. diubah menjadi H2O2, yang menyebabkan pembentukan radikal oksigen tambahan. Beberapa logam, seperti arsenit, secara tidak langsung menginduksi pembentukan ROS dengan aktivasi sistem penghasil radikal dalam sel
Arsenik adalah elemen yang sangat beracun yang menghasilkan berbagai ROS, termasuk superoksida (O2 2), oksigen singlet (1O2), radikal peroksil (ROO), oksida nitrat (NO), hidrogen peroksida (H2O2), dan radikal peroksil dimetilarsinik [( CH3) 2AsOO] .57 59 Senyawa arsen (III) dapat menghambat enzim antioksidan, terutama enzim yang bergantung pada GSH, seperti glutathione-S-transferases (GSTs), glutathione peroxidase (GSH-Px), dan GSH reduktase, melalui ikatan - masuk ke grup sulfhydryl ( SH) mereka.60,61
Timbal meningkatkan peroksidasi lipid.62 Penurunan aktivitas enzim SOD dan GPX otak yang signifikan terjadi setelah paparan timbal. 63,64 Penggantian seng, yang berfungsi sebagai kofaktor untuk banyak enzim oleh timbal, menyebabkan inaktivasi enzim tersebut. Paparan timbal dapat menyebabkan penghambatan GST dengan mempengaruhi tiol jaringan.
ROS yang dihasilkan oleh reaksi katalis logam dapat membentuk basis DNA. Tiga substitusi dasar, G / C, G / T, dan C / T, dapat terjadi sebagai akibat kerusakan oksidatif oleh ion logam, seperti Fe21, Cu21, dan Ni21. Reid et al65 menunjukkan bahwa G / C didominasi oleh Fe21 sedangkan substitusi C / T oleh Cu21 dan Ni21.
Antioksidan
Tubuh manusia dilengkapi dengan berbagai antioksidan yang berfungsi untuk mengimbangi efek oksidan. Untuk semua tujuan praktis, ini dapat dibagi menjadi kategori 2: enzimatik (Tabel 2) dan nonenzymatic (Tabel 3).
Enzimatik Antioksidan
Antioksidan enzimatik utama paru-paru adalah SOD (EC 1.15.1.11), katalase (EC 1.11.1.6), dan GSH-Px (EC 1.11.1.9). Selain enzim utama ini, antioksidan lainnya, termasuk heme oxygenase-1 (EC 1.14.99.3), dan protein redoks, seperti thioredoxins (TRXs, EC 1.8.4.10), peroxiredoxins (PRXs, EC 1.11.1.15), dan glutaredoxins, juga ditemukan pada memainkan peran penting dalam pertahanan antioksidan paru.
Karena superoksida adalah ROS utama yang dihasilkan dari berbagai sumber, penurunannya oleh SOD sangat penting untuk setiap sel. Semua bentuk 3 dari SOD, yaitu CuZn-SOD, Mn-SOD, dan EC-SOD, banyak diekspresikan di paru-paru manusia. Mn-SOD dilokalisasi dalam matriks mitokondria. EC-SOD terutama terlokalisasi dalam matriks ekstraselular, terutama di daerah-daerah yang mengandung serat kolagen tipe I yang tinggi dan sekitar pembuluh darah paru dan sistemik. Ini juga telah terdeteksi di epitel bronkial, epitel alveolar, dan makrofag alveolar.66,67 Secara keseluruhan, CuZn-SOD dan Mn-SOD pada umumnya dianggap bertindak sebagai pemulung massal radikal superoksida. Tingkat EC-SOD yang relatif tinggi di paru-paru dengan pengikatnya yang spesifik terhadap komponen matriks ekstraselular dapat mewakili komponen dasar proteksi matriks paru.68
H2O2 yang diproduksi oleh aksi SOD atau tindakan oksidase, seperti xanthine oxidase, dikurangi menjadi air dengan katalase dan GSH-Px. Katalase ada sebagai tetra- mer yang terdiri dari monomer identik 4, yang masing-masing mengandung gugus heme di tempat yang aktif. Degradasi H2O2 dilakukan melalui konversi antara konformasi 2 dengan katalase-ferricatalase (besi yang dikoordinasikan dengan air) dan senyawa I (besi yang dikomplekskan dengan atom oksigen). Katalase juga mengikat NADPH sebagai padanan pereduksi untuk mencegah inaktivasi oksidatif enzim (pembentukan senyawa II) oleh H2O2 karena direduksi menjadi air.69
Enzim dalam siklus redoks bertanggung jawab untuk pengurangan H2O2 dan hydroperoxides lipid (dihasilkan sebagai hasil peroksidasi lipid membran) termasuk GSH-Pxs.70 GSH-Pxs adalah keluarga enzim tetramer yang mengandung selenosistein asam amino unik di dalam situs aktif dan menggunakan tiol berberat molekul rendah, seperti GSH, untuk mengurangi H2O2 dan peroksida lipida ke alkohol yang sesuai. Empat GSH-Pxs telah dijelaskan, dikodekan oleh gen yang berbeda: GSH-Px-1 (GSH-Px) seluler ada di mana-mana dan mengurangi H2O2 dan peroksida asam lemak, namun tidak mengoksidasi lipid peroksil.71 Lipid yang diferensikan dikurangi dengan GSH yang terikat membran -Px-4 (phospholipid hydroperoxide GSH-Px), yang dapat menggunakan beberapa tiol berberat molekul rendah yang berbeda seperti mengurangi padanannya. GSH-Px-2 (gastrointestinal GSH-Px) dilokalisasi di sel epitel gastrointestinal di mana berfungsi untuk mengurangi peroksida diet.72 GSH-Px-3 (GSH-Px ekstraselular) adalah satu-satunya anggota keluarga GSH-Px yang tinggal di kompartemen ekstraselular dan diyakini sebagai salah satu enzim antioksidan ekstraselular terpenting pada mamalia. Dari jumlah tersebut, GSH-Px ekstraselular paling banyak diteliti di paru-paru manusia.73
Selain itu, pembuangan H2O2 terkait erat dengan beberapa enzim yang mengandung tiol, yaitu TRXs (TRX1 dan TRX2), reduktase tioredoksin (EC 1.8.1.9) (TRRs), PRXs (yang merupakan peroksidase tioredoksin), dan glutaredoksin.74
Dua TRXs dan TRRs telah dicirikan pada sel manusia, ada di sitosol dan mitokondria. Di paru-paru, TRX dan TRR diekspresikan dalam epitel bronkial dan alveolar dan makrofag. Enam PRX yang berbeda telah ditemukan di sel manusia, berbeda dalam kompartementalisasi ultrastruktural. Studi eksperimental telah mengungkapkan pentingnya PRX VI dalam melindungi epitel alveolar. Paru manusia mengungkapkan semua PRXs di epitel bronkial, epitel alveolar, dan makrofag.75 PRX V baru-baru ini ditemukan berfungsi sebagai peroxynitrite reductase, 76 yang berarti berfungsi sebagai senyawa pelindung potensial dalam pengembangan cedera paru yang dimediasi ROS. .77
Yang umum untuk antioksidan ini adalah kebutuhan NADPH sebagai ekuivalensi pereduksi. NADPH mempertahankan katalase dalam bentuk aktif dan digunakan sebagai kofaktor oleh TRX dan GSH reductase (EC 1.6.4.2), yang mengubah GSSG menjadi GSH, co-substrat untuk GSH-Pxs. NADPH intraselular, pada gilirannya, dihasilkan oleh pengurangan NADP1 oleh glukosa-6-fosfat dehidrogenase, enzim pengikatan pertama dan laju batas dari jalur fosfat pen- tose, selama konversi glukosa-6-fosfat menjadi 6-phosphogluconolactone. Dengan menghasilkan NADPH, glukosa-6-phosphate dehydrogenase adalah penentu penting kapasitas buffer GSH sitosolik (GSH / GSSG) dan oleh karena itu dapat dianggap sebagai enzim antioksidan penting dan penting. 78,79
GSTs (EC 2.5.1.18), keluarga enzim antioksidan lain, menonaktifkan metabolit sekunder, seperti aldehida tak jenuh, epoksida, dan hidroperoksida. Tiga keluarga utama GST telah dijelaskan: GST sitosol, GST mitokondria, 80,81 dan GST mikro terkait membran yang berperan dalam metabolisme eikosanoid dan GSH.82 Tujuh kelas GST sitosol diidentifikasi pada mamalia, yang disebut Alpha, Mu, Pi, Sigma, Theta, Omega, dan Zeta.83 86 Selama kondisi non-stres, kelas Mu dan Pi GSTs berinteraksi dengan kinase Ask1 dan JNK, masing-masing, dan menghambat kinase ini.87 89 Telah ditunjukkan bahwa GSTP1 terdisosiasi dari JNK sebagai respons terhadap stres oksidatif.89 GSTP1 juga berinteraksi secara fisik dengan PRX VI dan mengarah pada pemulihan aktivitas enzim PRX melalui glutationylation dari protein teroksidasi.90
Antioksidan nonenzymatik
Antioksidan nonenzimatik termasuk senyawa dengan berat molekul rendah, seperti vitamin (vitamin C dan E), b-karoten, asam urat, dan GSH, sebuah tripeptida (Lg-glutamyl-L-cysteinyl-L-glineine) yang terdiri dari tiol ( sulfhidril).
Vitamin C (Asam askorbat)
Vitamin C yang larut dalam air (asam askorbat) menyediakan kapasitas antioksidan fase berair intraselular dan ekstraselular terutama dengan mengais oksigen bebas oksigen. Ini mengubah vitamin E radikal bebas kembali ke vitamin E. Tingkat plasmanya telah terbukti menurun seiring bertambahnya usia.91,92
Vitamin E (a-tokoferol)
Vitamin E yang larut dalam lipid terkonsentrasi di situs interior hidrofobik membran sel dan merupakan pertahanan utama terhadap cedera membran akibat oksidan. Vitamin E menyumbangkan elektron ke radikal peroksil, yang diproduksi selama peroksidasi lipid. a-Tokoferol adalah bentuk paling aktif vitamin E dan antioksidan utama yang terikat pada sel. Vitamin E memicu apoptosis sel kanker dan menghambat formasi radikal bebas.93
Glutathione
GSH sangat melimpah di semua kompartemen sel dan merupakan antioksidan utama yang dapat larut. Rasio GSH / GSSG merupakan penentu utama stres oksidatif. GSH menunjukkan efek antioksidannya dalam beberapa cara. 94 Ini mendetoksifikasi peroksida hidrogen peroksida dan lipid melalui aksi GSH-Px. GSH menyumbangkan elektronnya ke H2O2 untuk menguranginya menjadi H2O dan O2. GSSG kembali diturunkan menjadi GSH oleh reduktase GSH yang menggunakan NAD (P) H sebagai donor elektron. GSH-Pxs juga penting untuk melindungi selaput sel dari peroksidasi lipid. Mengurangi glutathione menyumbangkan proton pada lipida membran dan melindungi mereka dari serangan oksidan.95
GSH adalah kofaktor untuk beberapa enzim detoksifikasi, seperti GSH-Px dan transferase. Ini memiliki peran dalam mengubah vitamin C dan E kembali ke bentuk aktifnya. GSH melindungi sel-sel melawan apoptosis dengan berinteraksi dengan jalur pensinyalan proapoptosis dan antiapoptotik. 94 Ini juga mengatur dan mengaktifkan beberapa faktor transkripsi, seperti AP-1, NF-kB, dan Sp-1.
Karotenoid (b-karoten)
Karotenoid adalah pigmen yang ditemukan pada tumbuhan. Terutama, b-karoten telah ditemukan bereaksi dengan radikal peroksil (ROO), hidroksil (OH), dan superoksida (O22.) .96 Karotenoid menunjukkan efek antioksidannya pada tekanan parsial oksigen rendah namun memiliki efek pro-oksidan pada oksigen yang lebih tinggi. konsentrasi.97 Baik karotenoid dan asam retinoat (RAs) mampu mengatur faktor transkripsi.98 b-Carotene menghambat aktivasi NF-kB yang diinduksi oleh oksidan dan interleukin (IL) -6 dan faktor nekrosis tumor - sebuah produksi. Karotenoid juga mempengaruhi apoptosis sel. Efek antiproliferatif RA telah ditunjukkan pada beberapa penelitian. Efek RA ini dimediasi terutama oleh reseptor asam retinoat dan bervariasi di antara jenis sel. Pada sel karsinoma mammae, reseptor asam retinoat ditunjukkan untuk memicu penghambatan pertumbuhan dengan menginduksi penangkapan sel siklus, apoptosis, atau keduanya. 99,100
PENGARUH STRES OKSIDATIF: MEKANISME GENETIK, FISIOLOGI, & BIOKIMIA
Stres oksidatif terjadi ketika keseimbangan antara antioksidan dan ROS terganggu karena penipisan antioksidan atau akumulasi ROS. Ketika stres oksidatif terjadi, sel mencoba untuk melawan efek oksidan dan mengembalikan keseimbangan redoks dengan mengaktifkan atau membungkam gen yang mengkode enzim defensif, faktor tranfiksi, dan protein struktural. Rasio 101,102 antara glutathione teroksidasi dan glukosa (2GSH / GSSG) adalah satu faktor penentu stres oksidatif yang penting dalam tubuh. Produksi ROS yang lebih tinggi dalam tubuh dapat mengubah struktur DNA, menghasilkan modifikasi protein dan lipid, pengaktifan beberapa faktor transkripsi yang disebabkan oleh stres, dan produksi sitokin pro-inflamasi dan anti-inflamasi.
Efek Stres Oksidatif Pada DNA
ROS dapat menyebabkan modifikasi DNA dengan beberapa cara, yang melibatkan degradasi basa, pemecahan DNA tunggal atau double-stranded, purin, pirimidin atau modifikasi terikat gula, mutasi, penghapusan atau translokasi, dan hubungan silang dengan protein. Sebagian besar modifikasi DNA (Gambar 1) sangat relevan untuk karsinogenesis, penuaan, dan penyakit neurodegeneratif, kardiovaskular, dan autoimun. Asap tembakau, logam redoks, dan logam nonredoks, seperti besi, kadmium, krom, dan arsenik, juga terlibat dalam karsinogenesis dan penuaan dengan menghasilkan radikal bebas atau ikatan dengan kelompok tiol. Pembentukan 8-OH-G adalah kerusakan DNA yang paling dikenal yang terjadi melalui stres oksidatif dan merupakan biomarker potensial untuk karsinogenesis.
Wilayah gen promotor berisi urutan konsensus untuk faktor transkripsi. Situs pengikat faktor transkripsi ini berisi urutan kaya GC yang rentan terhadap serangan oksidan. Pembentukan DNA 8-OH-G di situs pengikatan faktor transkripsi dapat memodifikasi pengikatan faktor transkripsi dan dengan demikian mengubah ekspresi gen terkait seperti yang telah ditunjukkan untuk urutan target AP-1 dan Sp-1 Selain 103-OH-G, 8 -cyclo-8,59 -deoxyadenosine (cyclo-dA) juga telah terbukti menghambat transkripsi dari gen reporter dalam sistem sel jika terletak di kotak TATA.29 Protein pengikat TATA memulai transkripsi dengan mengubah pembengkokan DNA . Pengikatan protein pengikat TATA mungkin terganggu oleh adanya cyclo-dA.
Stres oksidatif menyebabkan ketidakstabilan daerah mikrosatelit (daerah persendian pendek). Redoks ion logam aktif, radikal hidroksil meningkatkan ketidakstabilan mikrosatelit.105 Meskipun pemecahan DNA untai tunggal yang disebabkan oleh luka oksidan dapat ditoleransi dengan mudah oleh sel-sel, kerusakan DNA untai ganda yang disebabkan oleh radiasi pengion dapat menjadi ancaman signifikan bagi kelangsungan hidup sel. 106
Metilasi di pulau CpG dalam DNA adalah mekanisme epigenetik penting yang dapat menyebabkan pembungkaman gen. Oksidasi 5-MeCyt menjadi 5-hydroxymethyl uracil (5-OHMeUra) dapat terjadi melalui reaksi deaminasi / oksidasi intermediet timin atau 5-hydroxymethyl cytosine.107 Selain ekspresi gen modulasi, metilasi DNA juga tampaknya mempengaruhi organisasi kromatin.108 Pola metilasi DNA Aberrant yang disebabkan oleh serangan oksidatif juga mempengaruhi aktivitas perbaikan DNA.
Efek Stres Oksidatif Pada Lipid
ROS dapat menginduksi peroksidasi lipid dan mengganggu susunan lapisan ganda lipid membran yang dapat menonaktifkan reseptor dan enzim yang terikat membran dan meningkatkan permeabilitas jaringan.109 Produk peroksidasi lipid, seperti MDA dan aldehida tak jenuh, mampu menonaktifkan banyak protein seluler dengan membentuk persilangan protein -linkages.110 112 4-Hydroxy-2-nonenal menyebabkan penipisan GSH intraseluler dan menginduksi produksi peroksida, 113,114 mengaktifkan reseptor faktor pertumbuhan epidermal, 115 dan menginduksi produksi fibronektin.116 Produk peroksidasi lipid, seperti isoprostane dan zat reaktif asam thiobarbituric , telah digunakan sebagai biomarker tidak langsung dari stres oksidatif, dan peningkatan kadar ditunjukkan pada kondensat napas yang dihembuskan atau cairan lavage bronchoalveolar atau paru-paru pasien penyakit paru obstruktif kronik atau perokok.117-119
Efek Stres Oksidatif pada Protein
ROS dapat menyebabkan fragmentasi rantai peptida, perubahan muatan listrik protein, ikatan silang protein, dan oksidasi asam amino spesifik dan oleh karena itu menyebabkan peningkatan kerentanan terhadap proteolisis melalui degradasi oleh protease spesifik.120 Residu sistein dan metionin dalam protein adalah terutama lebih rentan terhadap oksidasi.121 Oksidasi gugus sulfhidril atau residu metionin protein menyebabkan perubahan konformasi, protein terungkap, dan degradasi.8,121-123 Enzim yang memiliki logam pada atau dekat dengan situs aktifnya sangat sensitif terhadap oksidasi katalis logam. Modifikasi oksidatif enzim telah terbukti menghambat aktivitasnya.124,125
Dalam beberapa kasus, oksidasi spesifik protein dapat terjadi. Sebagai contoh, metionin dapat dioksidasi metionin sulfoksida126 dan fenilalanin menjadi o-tirosin127; kelompok sulfhidril dapat dioksidasi untuk membentuk ikatan disulfida; gugus 128 dan karbonil dapat dimasukkan ke dalam rantai samping protein. Sinar gamma, oksidasi katalis logam, HOCl, dan ozon dapat menyebabkan pembentukan kelompok karbonil. 129
Efek Tegangan Oksidatif terhadap Transduksi Sinyal
ROS dapat menginduksi ekspresi beberapa gen yang terlibat dalam transduksi sinyal. 1,130 Rasio yang tinggi untuk GSH / GSSG penting untuk melindungi sel dari kerusakan oksidatif. Gangguan rasio ini menyebabkan aktivasi faktor transkripsi sensitif redoks, seperti NF-kB, AP-1, faktor inti sel T yang diaktifkan dan faktor 1 yang diinduksi hipoksia, yang terlibat dalam respons inflamasi. Aktivasi faktor transkripsi melalui ROS dicapai dengan kaskade transduksi sinyal yang mengirimkan informasi dari luar ke dalam sel. Reseptor tirosin kinase, sebagian besar reseptor faktor pertumbuhan, seperti reseptor faktor pertumbuhan epidermal, reseptor faktor pertumbuhan endotel vaskular, dan reseptor untuk faktor pertumbuhan yang diturunkan trombosit, protein tirosin fosfatase, dan serin / treonin kinase adalah target dari ROS.131-133 Kinase yang diatur sinyal ekstra-seluler, JNK, dan p38, yang merupakan anggota dari famili protein kinase yang diaktivasi oleh mitogen dan terlibat dalam beberapa proses dalam sel termasuk proliferasi, diferensiasi, dan apoptosis, juga dapat diatur oleh oksidan.
Di bawah kondisi stres oksidatif, residu sistein di situs pengikatan DNA c-Jun, beberapa subunit AP-1, dan penghambatan kB kinase mengalami S-glutathiolation yang dapat dibalik. Glutaredoxin dan TRX telah dilaporkan memainkan peran penting dalam regulasi jalur pensinyalan sensitif redoks, seperti NF-kB dan AP-1, p38 mitogen-activated protein kinase, dan JNK.134 137
NF-kB dapat diaktifkan sebagai respons terhadap kondisi stres oksidatif, seperti ROS, radikal bebas, dan penyinaran UV.138 Fosforilasi IkB membebaskan NF-kB dan memungkinkannya memasuki nukleus untuk mengaktifkan transkripsi gen.139 Sejumlah kinase memiliki telah dilaporkan memfosforilasi IkBs pada residu serin. Kinase ini adalah target sinyal oksidatif untuk aktivasi NF-kB.140 Agen pereduksi meningkatkan pengikatan DNA NF-kB, sedangkan agen pengoksidasi menghambat pengikatan DNA pada NF-kB. TRX dapat memberikan 2 tindakan yang berlawanan dalam regulasi NF-kB: di sitoplasma, ia memblokir degradasi IkB dan menghambat aktivasi NF-kB tetapi meningkatkan pengikatan DNA NF-kB dalam nukleus.141 Aktivasi NF-kB melalui degradasi terkait oksidasi IkB menghasilkan aktivasi beberapa gen yang berhubungan dengan pertahanan antioksidan. NF-kB mengatur ekspresi beberapa gen yang berpartisipasi dalam respon imun, seperti IL-1b, IL-6, tumor necrosis factor-a, IL-8, dan beberapa molekul adhesi.142,143 NF-kB juga mengatur angiogenesis dan proliferasi dan diferensiasi sel.
AP-1 juga diatur oleh negara redoks. Dengan adanya H2O2, beberapa ion logam dapat menginduksi aktivasi AP-1. Peningkatan rasio GSH / GSSG meningkatkan pengikatan AP-1 sementara GSSG menghambat pengikatan DNA pengikatan DNA AP-1.144 dari heterodimer Fos / Jun meningkat dengan pengurangan sistein tunggal yang dilestarikan dalam domain pengikat DNA masing-masing. protein, 145 sementara pengikatan DNA AP-1 dapat dihambat oleh GSSG pada banyak jenis sel, menunjukkan bahwa pembentukan ikatan disulfida oleh residu sistein menghambat pengikatan DNA AP-1.146,147 Transduksi sinyal melalui stres oksidatif dirangkum dalam Gambar 2.
KESIMPULAN
Stres oksidatif dapat timbul akibat kelebihan produksi ROS oleh reaksi metabolik yang menggunakan oksigen dan menggeser keseimbangan antara oksidan /antioksidan status yang mendukung oksidan. ROS diproduksi oleh aktivitas metabolik seluler dan faktor lingkungan, seperti polutan udara atau asap rokok. ROS adalah molekul yang sangat reaktif karena elektron yang tidak berpasangan dalam strukturnya dan bereaksi dengan beberapa makromolekul biologis dalam sel, seperti karbohidrat, asam nukleat, lipid, dan protein, dan mengubah fungsinya. ROS juga mempengaruhi ekspresi beberapa gen oleh upregulasi faktor transkripsi sensitif redoks dan pemodelan kromatin melalui perubahan asetilasi / deasetilasi histon. Peraturan negara redoks sangat penting untuk kelangsungan hidup sel, aktivasi, proliferasi, dan fungsi organ.
REFERENSI
1. Valko M, Rhodes CJ, Moncol J, Izakovic M, Mazur M. Radikal bebas, logam dan antioksidan pada kanker akibat stres oksidatif. Chem Biol Berinteraksi. 2006; 160: 1 40.
2. Halliwell B, Gutteridge JMC. Radikal Bebas di Biologi dan Kedokteran. 3rd ed. New York: Oxford University Press; 1999.
3. Marnett LJ. Kerusakan peroksidasi lemak dDNA oleh malondialdehida. Mutat Res. 1999; 424: 83 95.
4. Siems WG, Grune T, Esterbauer H. 4-Hydroxynonenal pembentukan selama iskemia dan reperfusi usus kecil tikus. �Ilmu Kehidupan. 1995;57:785-789.
5. ER. Stadtman. Peran spesies oksidan dalam penuaan. Curr Med Chem. 2004; 11: 1105 1112.
6. Wang MY, Dhingra K, Hittelman WN, Liehr JG, deAndrade M, Li DH. Aduk malondialdehida DNA putatif yang diinduksi peroksidasi lipid di jaringan payudara manusia. Cancer Epidemiol Biomarkers Sblm. 1996; 5: 705 710.
7. Jenner P. Stres oksidatif pada penyakit Parkinson. Ann Neurol. 2003; 53: S26 S36.
8. Lyras L, Cairns NJ, Jenner A, Jenner P, Halliwell B. Penilaian kerusakan oksidatif pada protein, lipid, dan DNA di otak dari pasien dengan penyakit Alzheimer. J Neurochem. 1997; 68: 2061 2069.
9. Sayre LM, Smith MA, Perry G. Kimia dan biokimia stres oksidatif pada penyakit neurodegeneratif. Curr Med Chem. 2001; 8: 721 738.
10. Toshniwal PK, Zarling EJ. Bukti peningkatan peroksidasi lipid pada multiple sclerosis. Res neurochem. 1992; 17: 205 207.
11. Dhalla NS, Temsah RM, Netticadan T. Peran stres oksidatif pada penyakit kardiovaskular. J Hypertens. 2000; 18: 655 673.
12. Kasparova S, Brezova V, Valko M, Horecky J, Mlynarik V, dkk. Studi stres oksidatif dalam model tikus hipoperfusi otak kronis. Neurochem Int. 2005; 46: 601 611.
13. Kerr S, Brosnan MJ, McIntyre M, Reid JL, Dominiczak AF, Hamilton CA. Produksi anion superoksida meningkat dalam model hipertensi genetik: peran endotelium. Hipertensi. 1999; 33: 1353 1358.
14. Kukreja RC, Hess ML. Sistem radikal bebas oksigen: dari persamaan melalui interaksi protein-membran hingga cedera dan perlindungan kardiovaskular. Cardiovasc Res. 1992; 26: 641 655.
15. Asami S, Manabe H, Miyake J, Tsurudome Y, Hirano T, dkk. Merokok menyebabkan peningkatan kerusakan DNA oksidatif, 8-hydroxydeoxyguanosine, di bagian tengah paru-paru manusia. Karsinogenesis. 1997; 18: 1763 1766.
16. Andreadis AA, Hazen SL, Comhair SA, Erzurum SC. Peristiwa oksidatif dan nitrosatif pada asma. Radic Bebas Berbagai Med. 2003; 35: 213 225.
17. Comhair SA, Ricci KS, Arroliga M, Lara AR, Dweik RA, dkk. Korelasi defisiensi superoksida dismutase sistemik dengan obstruksi aliran udara pada asma. Am J Respir Crit Perawatan Med. 2005; 172: 306 313.
18. Comhair SA, Xu W, Ghosh S, Thunnissen FB, Almasan A, dkk. Inaktivasi superoksida dismutase dalam patofisiologi renovasi dan reaktivitas jalan napas asma. Am J Pathol. 2005; 166: 663 674.
19. Dut R, Dizdar EA, Birben E, Sackesen C, Soyer OU, Besler T, Kalayci O. Stres oksidatif dan determinannya pada saluran udara anak dengan asma. Alergi. 2008; 63: 1605 1609.
20. Ercan H, Birben E, Dizdar EA, Keskin O, Karaaslan C, dkk. Stres oksidatif dan determinan genetik dan epidemiologi dari cedera oksidan pada asma masa kanak-kanak. J Alergi Clin Immunol. 2006; 118: 1097 1104.
21. Fitzpatrick AM, Teague WG, Holguin F, Yeh M, Brown LA. Program Penelitian Asma Parah. Homeostasis glutathione saluran napas diubah pada anak-anak dengan asma berat: bukti stres oksidan. J Alergi Clin Immunol. 2009; 123: 146 152.
22. Miller DM, Buettner GR, Aust SD. Logam transisi sebagai katalisator reaksi "autoksidasi". Radic Bebas Berbagai Med. 1990; 8: 95 108.
23. Dupuy C, Virion A, Ohayon R, Kaniewski J, D me D, Pommier J. Mekanisme pembentukan hidrogen peroksida yang dikatalisis oleh NADPH oksidase dalam membran plasma tiroid. J berbagai Chem. 1991; 266: 3739 3743.
24. Granger DN. Peran xantin oksidase dan granulosit pada cedera epfusi iskemia. Am J Physiol. 1988; 255: H1269 H1275.
25. Fenton HJH. Oksidasi asam tartarat dengan adanya besi. J Chem Soc. 1984; 65: 899 910.
26. Haber F, Weiss JJ. Dekomposisi katalitik hidrogen peroksida oleh garam besi. Proc R Soc Lond Ser A.1934; 147: 332 351.
27. Liochev SI, Fridovich I. The Haber Weiss bersepeda 70 tahun kemudian: pandangan alternatif. Redoks Rep.2002; 7: 55 57.
28. Klebanoff SJ. Myeloperoxidase: teman dan musuh. J Leukoc berbagai. 2005; 77: 598 625.
29. Whiteman M, Jenner A, Halliwell B. modifikasi basa yang diinduksi asam hipoklorit pada DNA timus betis yang diisolasi. Chem Res Toxicol. 1997; 10: 1240 1246.
30. Kulcharyk PA, Heinecke JW. Asam hipoklorit yang diproduksi oleh sistem myeloperoksidase fagosit manusia menginduksi hubungan silang kovalen antara DNA dan protein. Biokimia. 2001; 40: 3648 3656.
31. Brennan ML, Wu W, Fu X, Shen Z, Lagu W, dkk. Kisah dua kontroversi: mendefinisikan baik peran peroksidase dalam pembentukan nitrotyrosine in vivo menggunakan tikus yang kekurangan eosinofil peroksidase dan myeloperoksidase, dan sifat spesies nitrogen reaktif yang dihasilkan peroksidase. J berbagai Chem. 2002; 277: 17415 17427.
32. Denzler KL, Borchers MT, Crosby JR, Cieslewicz G, Hines EM, dkk. Degranulasi eosinofil ekstensif dan oksidasi protein saluran napas yang dimediasi peroksidase tidak terjadi pada model inflamasi paru tikus model ovalbumin-challenge. J Immunol. 2001; 167: 1672 1682.
33. van Dalen CJ, Winterbourn CC, Senthilmohan R, Kettle AJ. Nitrit sebagai substrat dan penghambat myeloperoksidase. Implikasi untuk produksi nitrasi dan asam hipoklorit di tempat peradangan. J berbagai Chem. 2000; 275: 11638 11644.
34. Kayu LG, Fitzgerald DA, Gibson PG, Cooper DM, Garg ML. Peroksidasi lipid yang ditentukan oleh isoprostan plasma berhubungan dengan keparahan penyakit pada asma ringan. Lemak. 2000; 35: 967 974.
35. Montuschi P, Corradi M, Ciabattoni G, Nightingale J, Kharitonov SA, Barnes PJ. Peningkatan 8-isoprostane, penanda stres oksidatif, pada kondensat yang dihembuskan pada pasien asma. Am J Respir Crit Perawatan Med. 1999; 160: 216 220.
36. Gereja DF, Pryor WA. Kimia radikal bebas dari asap rokok dan implikasi toksikologisnya. Perspektif Kesehatan Lingkungan. 1985; 64: 111 126.
37. Hiltermann JT, Lapperre TS, van Bree L, Steerenberg PA, Brahim JJ, dkk. Peradangan yang diinduksi ozon dinilai dalam sputum dan cairan lavage bronkial dari penderita asma: alat noninvasif baru dalam studi epidemiologi tentang polusi udara dan asma. Radic Bebas Berbagai Med. 1999; 27: 1448 1454.
38. Nightingale JA, Rogers DF, Barnes PJ. Pengaruh ozon yang dihirup pada oksida nitrat yang dihembuskan, fungsi paru, dan induksi sputum pada subjek normal dan penderita asma. Thorax. 1999; 54: 1061 1069.
39. Cho AK, Sioutas C, Miguel AH, Kumagai Y, Schmitz DA, dkk. Aktivitas redoks materi partikulat udara di berbagai lokasi di Cekungan Los Angeles. Res. Lingkungan 2005; 99: 40 47.
40. Comhair SA, Thomassen MJ, Erzurum SC. Induksi diferensial dari glutathione peroksidase ekstraseluler dan sintase oksida nitrat 2 dalam saluran udara orang sehat yang terpapar 100% O (2) atau asap rokok. Am J Respir Cell Mol berbagai. 2000; 23: 350 354.
41. Matthay MA, Geiser T, Matalon S, Ischiropoulos H. Cedera paru-paru yang dimediasi oksidan pada sindrom gangguan pernapasan akut. Crit Perawatan Med. 1999; 27: 2028 2030.
42. Biaglow JE, Mitchell JB, Diadakan K. Pentingnya peroksida dan superoksida dalam respons sinar-X. Int J Radiat Oncol Berbagai Phys. 1992; 22: 665 669.
43. Chiu SM, Xue LY, Friedman LR, Oleinick NL. Sensitisasi yang dimediasi ion tembaga dari situs lampiran matriks nuklir ke radiasi pengion. Biokimia. 1993; 32: 6214 6219.
44. Narayanan PK, Goodwin EH, Lehnert BE. Partikel alfa memulai produksi biologis anion superoksida dan hidrogen peroksida dalam sel manusia. Res kanker. 1997; 57: 3963 3971.
45. Tuttle SW, Varnes ME, Mitchell JB, Biaglow JE. Sensitivitas terhadap oksidan kimia dan radiasi dalam garis sel CHO kurang dalam aktivitas siklus pentosa oksidatif. Int J Radiat Oncol Berbagai Phys. 1992; 22: 671 675.
46. Guo G, Yan-Sanders Y, Lyn-Cook BD, Wang T, Tamae D, dkk. Mangan
Ekspresi gen superoksida dismutase dalam radiasi yang diinduksi
tanggapan adaptif. Mol Sel Berbagai. 2003; 23: 2362 2378.
47. Azzam EI, Toledo de Toledo, DR Spitz, JB Kecil. Metabolisme oksidatif
memodulasi transduksi sinyal dan pembentukan mikronukleus di pengamat
sel-sel dari fibroblas manusia normal yang diiradiasi dengan iradiasi. Kanker Res.
2002; 62: 5436 5442.
48. Leach JK, Van Tuyle G, Lin PS, Schmidt-Ullrich R, Mikkelsen RB.
Mengionisasi radiasi yang diinduksi, bergantung pada mitokondria dari reaktif
oksigen / nitrogen. Res kanker. 2001; 61: 3894 3901.
49. Dent P, Yacoub A, Fisher PB, Hagan MP, jalur Grant S. MAPK di
respons radiasi. Onkogen. 2003; 22: 5885 5896.
50. Wei SJ, Botero A, Hirota K, Bradbury CM, Markovina S, dkk. Thioredoxin
translokasi nuklir dan interaksi dengan faktor redoks-1 mengaktifkan faktor transkripsi AP-1 sebagai respons terhadap radiasi pengion. Res kanker. 2000; 60: 6688 6695.
51. Kadet J, Douki T, Gasparutto D, Ravanat JL. Kerusakan oksidatif pada DNA: pembentukan, pengukuran, dan fitur biokimia. Mutat Res. 2003; 531: 5 23.
52. Yokoya A, Cunniffe SM, O Neill P. Pengaruh hidrasi pada induksi putus untai dan lesi basa pada film DNA plasmid melalui gammaradiasi. J Am Chem Soc. 2002; 124: 8859 8866.
53. Janssen YM, Van Houten B, Borm PJ, Mossman BT. Respons sel dan jaringan terhadap kerusakan oksidatif. Investasi Lab. 1993; 69: 261 274.
54. Iwanaga M, Mori K, Iida T, Urata Y, Matsuo T, dkk. Faktor inti kappa B tergantung induksi sintetase gamma glutamylcysteine oleh radiasi pengion dalam sel glioblastoma manusia T98G. Radic Bebas Berbagai Med. 1998; 24: 1256 1268.
55. Stohs SJ, Bagchi D. Mekanisme oksidatif dalam toksisitas ion logam. Radic Bebas Berbagai Med. 1995; 18: 321 336.
56. Leonard SS, Harris GK, Shi X. Tegangan oksidatif yang diinduksi logam dan transduksi sinyal. Radic Bebas Berbagai Med. 2004; 37: 1921 1942.
57. Shi H, Shi X, Liu KJ. Mekanisme oksidatif toksisitas arsenik dan karsinogenesis. Mol Cell Biochem. 2004; 255: 67 78.
58. Pi J, Horiguchi S, Sun Y, Nikaido M, Shimojo N, Hayashi T. Mekanisme potensial untuk kerusakan pembentukan oksida nitrat yang disebabkan oleh paparan oral yang lama terhadap arsenat pada kelinci. Radik Bebas Berbagai Med. 2003; 35: 102 113.
59. Rin K, Kawaguchi K, Yamanaka K, Tezuka M, Oku N, Okada S. Tali DNA putus yang disebabkan oleh asam dimetilarsinat, suatu metabolit arsenik anorganik, diperkuat secara kuat oleh radikal anion superoksida. Biol Pharm Bull. 1995; 18: 45 58.
60. Anggota Parlemen Waalkes, Liu J, Ward JM, Diwan LA. Mekanisme yang mendasari karsinogenesis arsenik: hipersensitivitas mencit yang terpapar arsen anorganik selama gestasi. Toksikologi. 2004; 198: 31 38.
61. CM Schiller, Fowler BA, Woods JS. Pengaruh arsenik pada aktivasi dehidrogenase piruvat. Perspektif Kesehatan Lingkungan. 1977; 19: 205 207.
62. Monterio HP, Bechara EJH, Abdalla DSP. Keterlibatan radikal bebas dalam porfiria neurologis dan keracunan timbal. Mol Cell Biochem. 1991; 103: 73 83.
63. Tripathi RM, Raghunath R, Mahapatra S. Timbal darah dan pengaruhnya terhadap kadar Cd, Cu, Zn, Fe dan hemoglobin anak. Lingkungan Total Sci. 2001; 277: 161 168.
64. Nehru B, Dua R. Pengaruh selenium makanan pada neurotoksisitas timbal. J Lingkungan Pathol Toxicol Oncol. 1997; 16: 47 50.
65. Reid TM, Feig DI, Loeb LA. Mutagenesis oleh radikal oksigen yang diinduksi logam. Perspektif Kesehatan Lingkungan. 1994; 102 (suppl 3): 57 61.
66. Kinnula VL, Crapo JD. Superoksida dismutase di paru-paru dan penyakit paru-paru manusia. Am J Respir Crit Perawatan Med. 2003; 167: 1600 1619.
67. Kinnula VL. Produksi dan degradasi metabolit oksigen selama keadaan inflamasi di paru-paru manusia. Curr Drug Menargetkan Alergi Inflamasi. 2005; 4: 465 470.
68. Zelko IN, Mariani TJ, Folz RJ. Keluarga multigen superoksida dismutase: perbandingan struktur, evolusi, dan ekspresi gen CuZn-SOD (SOD1), Mn-SOD (SOD2), dan EC-SOD (SOD3). Radic Bebas Berbagai Med. 2002; 33: 337 349.
69. Kirkman HN, Rolfo M, Ferraris AM, Gaetani GF. Mekanisme proteksi katalase oleh NADPH. Kinetika dan stoikiometri. J berbagai Chem. 1999; 274: 13908 13914.
70. Floh L. Glutathione peroksidase. Dasar Kehidupan Sci. 1988; 49: 663 668.
71. Arthur JR. Glutathione peroksidase. Cell Mol Life Sci. 2000; 57: 1825 1835.
72. Chu FF, Doroshow JH, Esworthy RS. Ekspresi, karakterisasi, dan distribusi jaringan dari glutathione peroksidase yang bergantung pada selenium, GSHPx-GI. J berbagai Chem. 1993; 268: 2571 2576.
73. Comhair SA, Bhathena PR, Farver C, Thunnissen FB, Erzurum SC. Induksi glutathione peroksidase ekstraseluler di paru-paru asma: bukti regulasi redoks ekspresi dalam sel epitel saluran napas manusia. FASEB J.2001; 15: 70 78.
74. Gromer S, Urig S, Becker K. Sistem tioredoksin dari sains ke klinik. Med Res Rev.2004; 24: 40 89.
75. Kinnula VL, Lehtonen S, Kaarteenaho-Wiik R, Lakari E, P kk P, dkk. Ekspresi spesifik sel peroxiredoxin di paru-paru manusia dan sarcoidosis paru. Thorax. 2002; 57: 157 164.
76. Dubuisson M, Vander Stricht D, Clippe A, Etienne F, Nauser T, dkk. Peroxiredoxin 5 manusia adalah reduktase peroksinitrit. FEBS Lett. 2004; 571: 161 165.
77. Holmgren A. Fungsi antioksidan dari sistem tioredoksin dan glutaredoksin. Sinyal Redoks Antioksidan. 2000; 2: 811 820.
78. Dickinson DA, Forman HJ. Glutathione dalam pertahanan dan pensinyalan: pelajaran dari tiol kecil. Ann NY Acad Sci. 2002; 973: 488 504.
79. Sies H. Glutathione dan perannya dalam fungsi seluler. Radic Bebas Berbagai Med. 1999; 27: 916 921.
80. Ladner JE, Parsons JF, Rife CL, Gilliland GL, Armstrong RN. Jalur evolusi paralel untuk glutathione transferase: struktur dan mekanisme enzim kappa kelas mitokondria rGSTK1-1. Biokimia. 2004; 43: 52 61.
81. Robinson A, Huttley GA, Booth HS, Dewan PG. Pemodelan dan studi bioinformatika dari kelas kappa manusia glutathione transferase memprediksi keluarga transferase ketiga yang baru dengan homologi untuk isomerase 2-hidroksikromen-2-karboksilat prokariotik. Biochem J.2004; 379: 541 552.
82. Jakobsson PJ, Morgenstern R, Mancini J, Ford-Hutchinson A, Persson B. Ciri-ciri struktural umum MAPEGda superfamili yang tersebar luas dari membran terkait protein dengan fungsi yang sangat berbeda dalam metabolisme eicosanoid dan glutathione. Protein Sci. 1999; 8: 689 692.
83. Hayes JD, Pulford DJ. Keluarga supergen glutathione S-transferase: regulasi GST dan kontribusi isoenzim terhadap kemoproteksi kanker dan resistensi obat. Crit Rev Biochem Mol berbagai. 1995; 30: 445 600.
84. Armstrong RN. Struktur, mekanisme katalitik, dan evolusi transferase glutathione. Chem Res Toxicol. 1997; 10: 2 18.
85. Hayes JD, McLellan LI. Glutathione dan enzim yang bergantung pada glutathione mewakili pertahanan yang diatur secara terkoordinasi melawan stres oksidatif. Res Radic Bebas. 1999; 31: 273 300.
86. Sheehan D, Meade G, Foley VM, Dowd CA. Struktur, fungsi, dan evolusi transferase glutathione: implikasi untuk klasifikasi anggota non-mamalia dari superfamili enzim purba. Biochem J.2001; 360: 1 16.
87. Cho SG, Lee YH, Park HS, Ryoo K, Kang KW, dkk. Glutathione S-transferase Mu memodulasi sinyal yang diaktifkan stres dengan menekan kinase pengatur sinyal apoptosis 1. J Biol Chem. 2001; 276: 12749 12755.
88. Dorion S, Lambert H, Landry J. Aktivasi jalur pensinyalan p38 dengan kejutan panas melibatkan disosiasi glutathione S-transferase Mu dari Ask1. J berbagai Chem. 2002; 277: 30792 30797.
89. Adler V, Yin Z, Fuchs SY, Benezra M, Rosario L, dkk. Peraturan pensinyalan JNK oleh GSTp. EMBO J.1999; 18: 1321 1334.
90. Manevich Y, Feinstein SI, Fisher AB. Aktivasi enzim antioksidan peroksiredoksin 1-CYS memerlukan glutathionylation yang dimediasi oleh heterodimerisasi dengan pGST. Proc Natl Acad Sci US A.2004; 101: 3780 3785.
91. Bunker VW. Radikal bebas, antioksidan dan penuaan. Lab Medis Sci. 1992; 49: 299 312.
92. Mezzetti A, Lapenna D, Romano F, Costantini F, Pierdomenico SD, dkk. Stres oksidatif sistemik dan hubungannya dengan usia dan penyakit. J Am Geriatr Soc. 1996; 44: 823 827.
93. White E, Shannon JS, Patterson RE. Hubungan antara vitamin dan
penggunaan suplemen kalsium dan kanker usus besar. Cancer Epidemiol Biomarkers Sblm. 1997; 6: 769 774.
94. Masella R, Di Benedetto R, Vari R, Filesi C, Giovannini C. Mekanisme baru senyawa antioksidan alami dalam sistem biologis: keterlibatan glutathione dan enzim yang berhubungan dengan glutathione. J Nutr Biochem. 2005; 16: 577 586.
95. Curello S, Ceconi C, Bigoli C, Ferrari R, Albertini A, Guarnieri C. Perubahan status glutathione jantung setelah iskemia dan reperfusi. Experientia. 1985; 41: 42 43.
96. El-Agamey A, GM Lowe, DJ McGarvey, Mortensen A, Phillip DM, Truscott TG. Sifat kimiawi radikal karotenoid dan antioksidan / pro-oksidan. Arch Biochem Biophys. 2004; 430: 37 48.
97. Beras-Evans CA, Sampson J, Bramley PM, Holloway DE. Mengapa kita mengharapkan karotenoid menjadi antioksidan in vivo? Res Radic Bebas. 1997; 26: 381 398.
98. Niles RM. Jalur pensinyalan dalam kemoprevensi retinoid dan pengobatan kanker. Mutat Res. 2004; 555: 81 96.
99. Donato LJ, Noy N. Penekanan pertumbuhan karsinoma mammae oleh asam retinoat: gen proapoptosis adalah target untuk reseptor asam retinoat dan pensinyalan protein pengikat asam retinoat seluler II. Res kanker. 2005; 65: 8193 8199.
100. Niizuma H, Nakamura Y, Ozaki T, Nakanishi H, Ohira M, dkk. Bcl-2 adalah pengatur kunci untuk kematian sel apoptosis yang diinduksi asam retinoat pada neuroblastoma. Onkogen. 2006; 25: 5046 5055.
101. Dalton TP, Shertzer HG, Puga A. Pengaturan ekspresi gen oleh oksigen reaktif. Ann Rev Pharmacol Toxicol. 1999; 39: 67 101.
102. Scandalios JG. Respons genomik terhadap stres oksidatif. Dalam: Meyers RA, ed. Ensiklopedia Biologi Sel Molekuler dan Kedokteran Molekuler. Vol 5. 2nd ed. Weinheim, Jerman: Wiley-VCH; 2004: 489 512.
103. Ghosh R, Mitchell DL. Pengaruh kerusakan DNA oksidatif dalam elemen promotor pada pengikatan faktor transkripsi. Res asam nukleat. 1999; 27: 3213 3218.
104. Marietta C, Gulam H, Brooks PJ. Lesi tunggal 8, 50-cyclo-20-deoxyadenosine dalam kotak TATA mencegah pengikatan protein pengikat TATA dan sangat mengurangi transkripsi in vivo. Perbaikan DNA (Amst). 2002; 1: 967 975.
105. Jackson AL, Chen R, Loeb LA. Induksi ketidakstabilan mikrosatelit
oleh kerusakan DNA oksidatif. Proc Natl Acad Sci US A.1998; 95: 12468 12473.
106. Caldecott KW. Interaksi protein-protein selama perbaikan untai tunggal DNA mamalia. Biochem Soc Trans. 2003; 31: 247 251.
107. Cooke MS, Evans MD, Dizdaroglu M, Lunec J. Kerusakan DNA oksidatif: mekanisme, mutasi, dan penyakit. FASEB J.2003; 17: 1195 1214.
108. Jones PL, Wolffe AP. Hubungan antara organisasi kromatin dan metilasi DNA dalam menentukan ekspresi gen. Biol Kanker Semin. 1999; 9: 339 347.
109. Girotti AW. Mekanisme peroksidasi lipid. J Gratis Radic Berbagai Med. 1985; 1: 87 95.
110. Siu GM, Draper HH. Metabolisme malonaldehyde in vivo dan in vitro. Lemak. 1982; 17: 349 355.
111. Esterbauer H, Koller E, Lengan RG, Koster JF. Kemungkinan keterlibatan produk peroksidasi lipid 4-hydroxynonenal dalam pembentukan kromolipid fluoresen. Biochem J. 1986; 239: 405 409.
112. Hagihara M, Nishigaki I, Maseki M, Yagi K. Perubahan tergantung usia pada tingkat lipid peroksida dalam fraksi lipoprotein serum manusia. J Gerontol. 1984; 39: 269 272.
113. Keller JN, Mark RJ, Bruce AJ, Blanc E, Rothstein JD, dkk. 4- Hidroksinonenal, produk aldehid dari peroksidasi lipid membran, mengganggu transportasi glutamat dan fungsi mitokondria di sinaptosom. Ilmu saraf. 1997; 806: 85 96.
114. Uchida K, Shiraishi M, Naito Y, Torii Y, Nakamura Y, Osawa T. Aktivasi jalur pensinyalan stres oleh produk akhir peroksidasi lipid. 4-hidroksi-2-nonenal adalah penginduksi potensial produksi peroksida intraseluler. J berbagai Chem. 1999; 274: 2234 2242.
115. Suc I, Meilhac O, Lajoie-Mazenc I, Vandaele J, Jurgens G, Salvayre R, Negre-Salvayre A. Aktivasi reseptor EGF oleh LDL teroksidasi. FASEB J.1998; 12: 665 671.
116. Tsukagoshi H, Kawata T, Shimizu Y, Ishizuka T, Dobashi K, Mori M. 4-Hydroxy-2-nonenal meningkatkan produksi fibronektin oleh IMR-90 fibroblas paru-paru manusia sebagian melalui aktivasi sinyal ekstraseluler terkait reseptor faktor pertumbuhan epidermal- jalur kinase p44 / 42 yang diatur. Toxicol Appl Pharmacol. 2002; 184: 127 135.
117. Montuschi P, Collins JV, Ciabattoni G, Lazzeri N, Corradi M, Kharitonov SA, Barnes PJ. Menghembuskan 8-isoprostane sebagai biomarker in vivo stres oksidatif paru pada pasien PPOK dan perokok sehat. Am J Respir Crit Perawatan Med. 2000; 162: 1175 1177.
118. Morrison D, Rahman I, Lannan S, MacNee W. Permeabilitas epitel, peradangan, dan stres oksidan di ruang udara perokok. Am J Respir Crit Perawatan Med. 1999; 159: 473 479.
119. Nowak D, Kasielski M, Antczak A, Pietras T, Bialasiewicz P. Peningkatan kandungan zat reaktif asam thiobarbituric dan hidrogen peroksida dalam kondensat nafas kadaluwarsa pasien dengan penyakit paru obstruktif kronik stabil: tidak ada efek signifikan dari merokok. Respir Med. 1999; 93: 389 396.
120. Kelly FJ, Mudway IS. Oksidasi protein di antarmuka udara-paru. Asam amino. 2003; 25: 375 396.
121. Dean RT, Roberts CR, Jessup W. Fragmentasi polipeptida ekstraseluler dan intraseluler oleh radikal bebas. Prog Clin berbagai Res. 1985; 180: 341 350.
122. Keck RG. Penggunaan t-butil hidroperoksida sebagai probe untuk oksidasi metionin dalam protein. Biokem Anal. 1996; 236: 56 62.
123. Davies KJ. Kerusakan dan degradasi protein oleh radikal oksigen. I. Aspek umum. J berbagai Chem. 1987; 262: 9895 9901.
124. Stadtman ER. Oksidasi katalis ion logam dari protein: mekanisme biokimia dan konsekuensi biologis. Radic Biol Med gratis.
1990; 9: 315 325.
125. Fucci L, Oliver CN, Coon MJ, Stadtman ER. Inaktivasi enzim metabolik utama melalui reaksi oksidasi fungsi campuran: kemungkinan berimplikasi pada pergantian protein dan penuaan. Proc Natl Acad Sci US A. 1983; 80: 1521 1525.
126. Stadtman ER, Moskovitz J, Levine RL. Oksidasi residu metionin protein: konsekuensi biologis. Sinyal Redoks Antioksidan. 2003; 5: 577 582.
127. Stadtman ER, Levine RL. Oksidasi asam amino bebas dan residu asam amino dalam protein yang dimediasi radikal bebas. Asam amino. 2003; 25: 207 218.
128. Stadtman ER. Oksidasi protein dalam penuaan dan penyakit terkait usia. Ann NY Acad Sci. 2001; 928: 22 38.
129. Shacter E. Kuantifikasi dan signifikansi oksidasi protein dalam sampel biologis. Drug Metab Rev.2000; 32: 307 326.
130. Poli G, Leonarduzzi G, Biasi F, Chiarpotto E. Stres oksidatif dan pensinyalan sel. Curr Med Chem. 2004; 11: 1163 1182.
131. Neufeld G, Cohen T, Gengrinovitch S, Poltorak Z. Faktor pertumbuhan endotel vaskular (VEGF) dan reseptornya. FASEB J.1999; 13: 9 22.
132. Sundaresan M, Yu ZX, Ferrans VJ, Sulciner DJ, Gutkind JS, dkk. Pengaturan generasi spesies oksigen reaktif dalam fibroblas oleh Rac1. Biochem J. 1996; 318: 379 382.
133. Sun T, Oberley LW. Regulasi redoks dari aktivator transkripsi. Radic Bebas Berbagai Med. 1996; 21: 335 348.
134. Klatt P, Molina EP, De Lacoba MG, Padilla CA, Martinez-Galesteo E, Barcena JA, Lamas S. Redoks regulasi pengikatan DNA c-Jun dengan S-glutathiolation reversibel. FASEB J.1999; 13: 1481 1490.
135. Reynaert NL, Ckless K, Guala AS, Wouters EF, van der Vliet A, Janssen Heininger
YM. Deteksi in situ protein S-glutathionylated setelah turunan sistein katalis glutaredoxin-1. Biochim Biophys Acta. 2006; 1760: 380 387.
136. Reynaert NL, Wouters EF, Janssen-Heininger YM. Modulasi glutaredoxin-1
ekspresi dalam model tikus penyakit saluran napas alergi. Am J Respir Cell Mol berbagai. 2007; 36: 147 151.
137. Filomeni G, Rotilio G, Ciriolo MR. Pensinyalan sel dan sistem redoks glutathione. Biochem Pharmacol. 2002; 64: 1057 1064.
138. Pande V, Ramos MJ. Pengenalan molekuler dari 15-deoxydelta (12,14) prostaglandin J (2) oleh faktor inti-kappa B dan protein seluler lainnya. Bioorg Med Chem Lett. 2005; 15: 4057 4063.
139. Perkins ND. Mengintegrasikan jalur pensinyalan sel dengan fungsi NF-kappaB dan IKK. Nat Rev Mol Sel Berbagai. 2007; 8: 49 62.
140. Gilmore TD. Pengantar NF-kappaB: pemain, jalur, perspektif. Onkogen. 2006; 25: 6680 6684.
141. Hirota K, Murata M, Sachi Y, Nakamura H, Takeuchi J, Mori K, Yodoi J. Peran berbeda dari tioredoksin dalam sitoplasma dan di dalam nukleus. Mekanisme dua langkah regulasi redoks faktor transkripsi NF-kappaB. J berbagai Chem. 1999; 274: 27891 27897.
142. Lingkungan PA. Peran komplemen, kemokin dan sitokin regulasi pada cedera paru akut. Ann NY Acad Sci. 1996; 796: 104 112.
143. Akira S, Kishimoto A. NF-IL6 dan NF-kB dalam regulasi gen sitokin. Adv Immunol. 1997; 65: 1 46.
144. Meyer M, Schreck R, Baeuerle PA. H2O2 dan antioksidan memiliki efek berlawanan pada aktivasi NF-kappa B dan AP-1 dalam sel utuh: AP-1 sebagai faktor responsif antioksidan sekunder. EMBO J.1993; 12: 2005 2015.
145. Abate C, Patel L, Rausher FJ, Curran T. Redoks regulasi aktivitas pengikatan DNA fos dan jun secara in vitro. Ilmu. 1990; 249: 1157 1161.
146. Galter D, Mihm S, Droge W. Efek berbeda dari glutathione disulphide pada faktor transkripsi inti kB dan aktivator protein-1. Eur J Biochem. 1994; 221: 639 648.
147. Aktivitas transkripsi Hirota K, Matsui M, Iwata S, Nishiyama A, Mori K, Yodoi J. AP-1 diatur oleh hubungan langsung antara tioredoksin dan Ref-1. Proc Natl Acad Sci US A.1997; 94: 3633 3638.
Alat Temukan Praktisi IFM adalah jaringan rujukan terbesar dalam Pengobatan Fungsional, dibuat untuk membantu pasien menemukan praktisi Pengobatan Fungsional di mana pun di dunia. Praktisi Bersertifikat IFM terdaftar pertama dalam hasil pencarian, mengingat pendidikan ekstensif mereka dalam Kedokteran Fungsional