Back Clinic Tim Chiropractic dan Pengobatan Fungsional Anti Aging. Tubuh kita berada dalam pertempuran yang konstan dan tidak pernah berakhir untuk bertahan hidup. Sel dilahirkan, sel dihancurkan. Para ilmuwan memperkirakan bahwa setiap sel harus menahan lebih dari 10,000 serangan individu dari spesies oksigen reaktif (ROS) atau radikal bebas. Tanpa Gagal, tubuh memiliki sistem penyembuhan diri yang luar biasa yang menahan serangan dan membangun kembali apa yang telah rusak atau hancur. Inilah keindahan desain kami.
Untuk memahami biologi penuaan dan menerjemahkan wawasan ilmiah ke dalam intervensi yang meningkatkan kesehatan usia lanjut melalui perawatan. Sangat berguna untuk memiliki pandangan konsensus yang jelas tentang apa sebenarnya yang dimaksud dengan perawatan anti-penuaan.
Sejak sebelum masa pencarian Ponce de Leon untuk umur panjang, manusia selalu terpikat oleh kesempatan awet muda. Perawatan kiropraktik dengan gerakan kesehatannya adalah metode yang ampuh untuk menstabilkan dan meningkatkan kemampuan penyembuhan diri ini. Dr Alex Jimenez membahas konsep seputar pandora anti-penuaan.
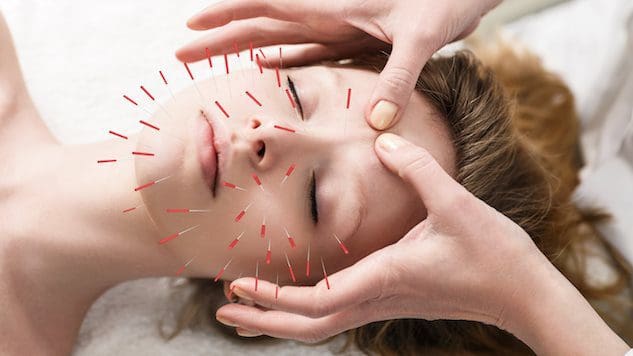
Bagi individu yang ingin meningkatkan atau menjaga kesehatan kulit, apakah penggunaan akupunktur dapat membantu memperbaiki kulit dan melawan proses penuaan?
Akupunktur Kosmetik
Akupunktur kosmetik mengikuti praktik akupunktur tradisional dengan memasukkan jarum. Tujuannya adalah untuk membalikkan tanda-tanda penuaan dan meningkatkan kesehatan kulit. Kadang-kadang disebut sebagai peremajaan wajah akupunktur, yang telah digunakan sebagai alternatif dari bedah pengencangan wajah dan prosedur konvensional lainnya. Studi pendahuluan telah meneliti bagaimana hal ini dapat membantu menghilangkan bintik-bintik penuaan, mengangkat kelopak mata yang kendur, dan mengurangi kerutan. (Younghee Yun dkk., 2013)
Bagaimana Akupunktur Bekerja
Dalam pengobatan tradisional Tiongkok atau TCM, akupunktur telah lama digunakan untuk meningkatkan aliran energi – qi atau chi – ke seluruh tubuh. Energi ini diyakini bersirkulasi melalui jalur energi yang disebut meridian. Ketika terjadi gangguan kesehatan, menurut TCM, ada hambatan atau penyumbatan pada sirkulasi.
Ahli akupunktur dapat memulihkan sirkulasi/aliran optimal dan meningkatkan kesehatan dengan menusukkan jarum ke titik akupuntur tertentu. (Institut Kesehatan Nasional, 2007)
Akupunktur Kosmetik
Akupunktur kosmetik dikatakan dapat meningkatkan kesehatan kulit dan bertindak sebagai pengobatan anti penuaan dengan merangsang produksi kolagen. Protein ini merupakan komponen utama kulit. Lapisan dalam kulit kehilangan kolagen dan kekencangannya seiring bertambahnya usia. Namun, penelitian lebih lanjut diperlukan untuk mendukung klaim bahwa akupunktur dapat meningkatkan produksi kolagen. Beberapa orang menyarankan akupunktur kosmetik membantu meremajakan kulit dengan meningkatkan energi tubuh secara keseluruhan. Sebuah penelitian menemukan bahwa individu melihat peningkatan setelah lima sesi akupunktur kosmetik wajah. (Younghee Yun dkk., 2013) Namun, disarankan agar sepuluh perawatan dilakukan sekali atau dua kali seminggu untuk hasil yang optimal. Setelah itu, perawatan pemeliharaan dilakukan setiap empat hingga delapan minggu. Berbeda dengan Botox atau dermal filler, akupunktur kosmetik bukanlah solusi yang cepat. Fokusnya adalah menciptakan perubahan jangka panjang pada kulit dan tubuh, yang berarti perbaikan:
Ketika jarum ditusukkan ke dalam kulit, akan timbul luka yang disebut mikrotrauma positif. Kemampuan penyembuhan dan perbaikan alami tubuh aktif ketika merasakan luka-luka ini. Tusukan ini merangsang sistem limfatik dan peredaran darah, yang mengantarkan nutrisi dan oksigen ke sel-sel kulit, memberi nutrisi dari dalam ke luar.
Ini membantu meratakan warna kulit dan membuat kulit bersinar.
Mikrotrauma positif juga merangsang produksi kolagen.
Ini membantu meningkatkan elastisitas, meminimalkan garis dan kerutan.
Alternatif
Beberapa pengobatan alami dapat membantu meningkatkan kesehatan kulit dan menawarkan manfaat anti penuaan. Ceramide adalah molekul lemak yang ditemukan secara alami di lapisan atas kulit dan merupakan bahan yang digunakan dalam produk perawatan kulit. Ini dapat melindungi kulit dari kekeringan akibat penuaan. (L Di Marzio 2008) Penelitian awal menunjukkan bahwa mengoleskan teh putih pada kulit dapat melawan pemecahan kolagen dan elastin – protein yang mendukung elastisitas kulit dan mencegah kendur). Ada juga bukti bahwa bahan alami seperti minyak argan, minyak borage, dan seabuckthorn dapat memberikan manfaat pelembab yang dapat memperbaiki kulit.(Tamsyn SA Thring dkk., 2009)
Meskipun bukti lebih lanjut tentang akupunktur kosmetik diperlukan, mengintegrasikan akupunktur dapat membantu mengelola stres dan meningkatkan kesehatan secara keseluruhan. Individu yang mempertimbangkan akupunktur kosmetik harus berkonsultasi dengan penyedia layanan kesehatan utama mereka untuk mengetahui apakah itu tepat untuk mereka.
Meningkatkan Kesehatan Bersama: Merangkul Evaluasi dan Perawatan Multidisiplin
Referensi
Yun, Y., Kim, S., Kim, M., Kim, K., Park, JS, & Choi, I. (2013). Pengaruh akupunktur kosmetik wajah pada elastisitas wajah: studi percontohan berlabel terbuka dan satu lengan. Pengobatan komplementer dan alternatif berbasis bukti : eCAM, 2013, 424313. doi.org/10.1155/2013/424313
Pusat Nasional untuk Pengobatan Komplementer dan Alternatif. (2007). Akupunktur: Sebuah Pengantar. Situs Web Pusat Nasional untuk Pengobatan Komplementer dan Alternatif. choimd.com/downloads/NIH-info-on-akupunktur.pdf
Kuge, H., Mori, H., Tanaka, TH, & Tsuji, R. (2021). Keandalan dan Validitas Lembar Periksa Wajah (FCS): Daftar Periksa Kepuasan Diri dengan Akupunktur Kosmetik. Obat-obatan (Basel, Swiss), 8(4), 18. doi.org/10.3390/medicines8040018
Di Marzio, L., Cinque, B., Cupelli, F., De Simone, C., Cifone, MG, & Giuliani, M. (2008). Peningkatan kadar ceramide kulit pada subjek lanjut usia setelah aplikasi topikal jangka pendek dari bakteri sphingomyelinase dari Streptococcus thermophilus. Jurnal internasional imunopatologi dan farmakologi, 21(1), 137–143. doi.org/10.1177/039463200802100115
Thring, TS, Hili, P., & Naughton, DP (2009). Aktivitas anti-kolagenase, anti-elastase dan anti-oksidan dari ekstrak 21 tanaman. Pengobatan komplementer dan alternatif BMC, 9, 27. doi.org/10.1186/1472-6882-9-27
Menjaga tulang belakang seseorang dalam kondisi prima sama dengan mengurangi rasa sakit dan lebih banyak mobilitas, fleksibilitas, dan kebebasan. Tubuh menjadi lelah dan merupakan efek alami dari penuaan yang terjadi pada kita semua. Masalah tulang belakang yang terkait dengan penuaan bisa menjadi serius jika tidak ditangani dan ditindaklanjuti dengan latihan, peregangan, dan perawatan kiropraktik.
Penuaan dan Punggung
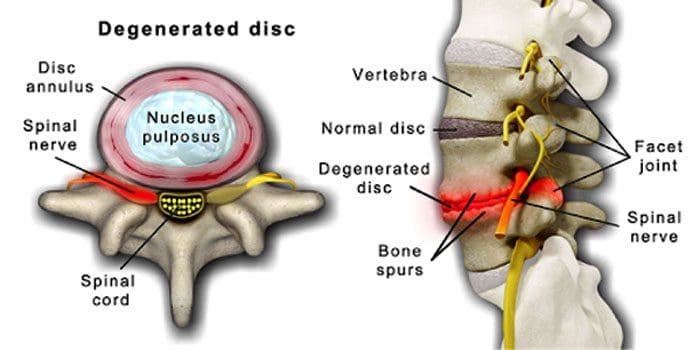
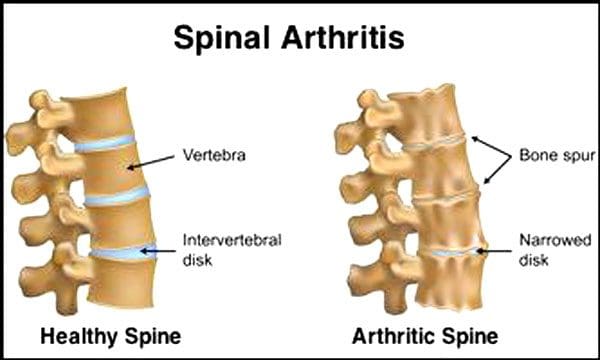
Kerusakan cakram dan sendi tulang belakang adalah hal yang normal seiring bertambahnya usia. Stenosis tulang belakang atau penyempitan saluran tulang belakang juga bisa menjadi bagian dari proses penuaan. Dua kondisi yang disebabkan oleh penuaan adalah penyakit degeneratif dan radang sendi itu juga bisa termasuk pengerasan ligamen tulang belakang dan osteoporosis.
Penyakit cakram degeneratif dialami oleh 40% individu yang berusia 40 tahun
Meningkat menjadi 80% untuk individu berusia 80 tahun ke atas.
Itu berpusat di sekitar cakram yang berangsur-angsur berubah dari sebagian besar air menjadi sebagian besar lemak.
Jika sudah gemuk, cakram menjadi menyempit dan kehilangan elastisitasnya.
Pusat Pengendalian dan Pencegahan Penyakit mengatakan itu 23% orang dewasa Amerika menderita radang sendi. Ini adalah kondisi yang terutama memengaruhi sendi facet. Sendi menjadi bengkak, yang mengurangi rentang gerak dan dapat mengenai saraf tulang belakang, menyebabkan nyeri, kelemahan, dan linu panggul. Seiring waktu, ligamen di sekitar dan di tulang belakang menegang, mengurangi rentang gerak, menyebabkan stenosis. Keropos tulang, atau osteoporosis, disebabkan oleh perubahan hormon dan faktor lain seperti nutrisi. Penuaan adalah proses alami, tetapi individu dapat membantu tulang punggung mereka tetap dalam kondisi prima berapa pun usia mereka.
Mempraktikkan Postur Tubuh yang Sehat
Langsung saja mekanisme tubuh sehat yang tepat Adalah sebuah keharusan. Menjaga dan memperhatikan postur tubuh menjaga keselarasan dan menjaga keseimbangan tubuh. Postur tubuh yang sehat akan membantu mengurangi efek dari:
Stenosis spinal
Penyakit degeneratif
Herniasi
Risiko patah tulang belakang
Mempraktikkan postur tubuh yang benar meliputi:
Kurangi membungkuk
Pastikan workstation dalam kondisi terbaik dan suara ergonomis
Pastikan untuk menekuk lutut saat mengangkat dan jaga agar tulang belakang se-vertikal mungkin.
Yoga
Yoga dapat sangat bermanfaat untuk tulang belakang yang lebih sehat dan lebih muda. Yoga memenuhi tiga area untuk menjaga tulang belakang tetap dalam kondisi prima. Ini termasuk:
Latihan rutin
Menjaga fleksibilitas
Mencapai berat badan ideal
Yoga adalah aktivitas tulang belakang yang menentang usia. Karena itu:
Mempertahankan kekuatan
keluwesan
Sikap
Saldo
Dapat membantu untuk berbagai kondisi tulang belakang, khususnya nyeri artritis
Jatuh dapat menyebabkan cedera serius. Yoga juga dapat membantu melatih keseimbangan.
Temui Chiropractor
Pengobatan preventif adalah kunci untuk menjaga tubuh tetap sehat, awet muda, dan sekuat mungkin. Pemeriksaan chiropractic dapat menentukan apakah ada masalah tulang belakang dan diagnosis untuk mengembangkan rencana perawatan yang optimal. Jika fungsi tubuh terbatas karena nyeri di punggung dan / atau tungkai, hubungi Cedera Medical Chiropractic dan Klinik Pengobatan Fungsional dan dapatkan tulang belakang kembali ke kondisi semula.
Komposisi tubuh
Latihan / Stabilitas Bola Curl
Latihan ini bekerja pada kelompok otot khusus untuk kekuatan tulang belakang dan meliputi:
Paha belakang
Otot bokong
Perut dalam
Penculik dan rotator pinggul
Latihan seperti ini adalah salah satu cara paling efektif untuk membangun kekuatan dan ketahanan fungsional pada paha belakang, pinggul, dan mencegah cedera. Untuk melakukan latihan ini:
Berbaring telentang dengan lutut ditekuk
Angkat kaki sehingga bagian bawah kaki bertumpu pada bola latihan
Gulingkan kaki Anda hingga lurus
Tahan posisi itu selama satu atau dua detik
Kembali ke gerakan puncak sambil meremas paha belakang
Mengerjakan otot-otot ini akan membantu mempermudah gerakan jongkok, menerjang, atau menekuk pada tulang belakang.
Penafian Entri Blog Dr. Alex Jimenez
Cakupan informasi kami terbatas pada chiropraktik, muskuloskeletal, obat-obatan fisik, kesehatan, dan masalah kesehatan sensitif dan / atau artikel, topik, dan diskusi kedokteran fungsional. Kami menggunakan protokol kesehatan & kebugaran fungsional untuk merawat dan mendukung perawatan cedera atau gangguan pada sistem muskuloskeletal. Postingan, topik, subjek, dan wawasan kami mencakup masalah klinis, masalah, dan topik yang terkait dan mendukung secara langsung atau tidak langsung ruang lingkup praktik klinis kami. *
Kantor kami telah melakukan upaya yang wajar untuk memberikan kutipan yang mendukung dan telah mengidentifikasi studi penelitian yang relevan atau studi yang mendukung posting kami. Kami juga menyediakan salinan studi penelitian pendukung untuk dewan dan atau publik atas permintaan. Kami memahami bahwa kami mencakup hal-hal yang memerlukan penjelasan tambahan tentang bagaimana hal itu dapat membantu dalam rencana perawatan atau protokol perawatan tertentu; Oleh karena itu, untuk membahas lebih lanjut pokok bahasan di atas, silakan bertanya kepada Dr. Alex Jimenez atau hubungi kami di 915-850-0900. Penyedia Berlisensi di Texas & New Mexico *
Makanan yang kita makan dapat berpotensi bermanfaat atau berbahaya bagi kesehatan kita. Nutrisi yang buruk dapat menyebabkan berbagai masalah kesehatan, termasuk obesitas, penyakit kardiovaskular, dan diabetes tipe 2. Sementara itu, nutrisi yang tepat dapat membuat Anda merasa berenergi, mengurangi risiko masalah kesehatan, serta membantu menjaga dan mengatur berat badan yang sehat. Jika Anda ingin mempromosikan umur panjang, Anda harus mengisi tubuh Anda dengan makanan yang baik. Dalam artikel berikut, kami akan mencantumkan beberapa makanan enak yang pada akhirnya dapat membantu mempromosikan umur panjang dengan juga membantu meningkatkan kesehatan dan kesejahteraan secara keseluruhan.
Sayuran menyalak
Sayuran kucifer memiliki kemampuan unik untuk mengubah hormon kita, memicu sistem detoksifikasi alami tubuh, dan bahkan mengurangi pertumbuhan sel kanker. Ini harus dikunyah seluruhnya atau dimakan diparut, dicincang, dijus, atau diblender untuk melepaskan khasiatnya yang bermanfaat. Sulforaphane, yang ditemukan dalam sayuran silangan, juga telah ditemukan untuk membantu melindungi dinding pembuluh darah dari peradangan yang dapat menyebabkan penyakit jantung. Sayuran kucifer, seperti kangkung, kubis, kubis Brussel, kembang kol, dan brokoli adalah beberapa makanan padat nutrisi di dunia.
Salad hijau
Sayuran hijau mentah memiliki kurang dari 100 kalori per pon, yang menjadikannya makanan yang sempurna untuk menurunkan berat badan. Makan lebih banyak salad hijau juga telah dikaitkan dengan penurunan risiko serangan jantung, stroke, diabetes, dan beberapa jenis kanker. Sayuran hijau mentah juga kaya akan vitamin B esensial, ditambah lutein dan zeaxanthin, karoten yang dapat membantu melindungi mata. Fitokimia yang larut dalam lemak, seperti karoten, ditemukan dalam salad sayuran seperti selada, bayam, kangkung, sawi hijau, dan sawi juga memiliki efek antioksidan dan antiinflamasi dalam tubuh.
Kacang-kacangan
Kacang-kacangan adalah makanan rendah glikemik dan sumber lemak sehat, protein nabati, serat, antioksidan, pitosterol, dan mineral, yang juga membantu mengurangi beban glikemik dari seluruh makanan, menjadikannya bagian penting dari anti-diabetes diet. Terlepas dari kepadatan kalori mereka, makan kacang dapat membantu menurunkan berat badan. Kacang juga dapat mengurangi kolesterol dan membantu mengurangi risiko penyakit jantung.
Benih
Biji-bijian, seperti kacang-kacangan, juga menyediakan lemak, antioksidan, dan mineral yang sehat, namun memiliki lebih banyak protein dan kaya akan mineral. Biji chia, rami, dan rami kaya akan lemak omega-3. Chia, rami, dan biji wijen juga kaya lignan atau fitoestrogen yang melawan kanker payudara. Apalagi biji wijen kaya akan kalsium dan vitamin E, dan biji labu kaya akan seng.
Berries
Buah beri adalah buah yang kaya antioksidan yang dapat membantu meningkatkan kesehatan jantung. Studi penelitian di mana partisipan makan stroberi atau blueberry setiap hari selama beberapa minggu melaporkan peningkatan tekanan darah, kolesterol total dan LDL, dan bahkan tanda-tanda stres oksidatif. Berry juga memiliki sifat anti-kanker dan telah terbukti membantu mencegah penurunan kognitif yang terkait dengan penuaan.
Delima
Fitokimia yang paling terkenal dalam buah delima, punicalagin, bertanggung jawab atas lebih dari setengah aktivitas antioksidan buah. Fitokimia buah delima memiliki manfaat anti kanker, pelindung jantung, dan kesehatan otak. Dalam satu studi penelitian, orang dewasa yang lebih tua yang minum jus delima setiap hari selama 28 hari tampil lebih baik pada tes memori dibandingkan dengan mereka yang minum minuman plasebo.
Kacang
Mengonsumsi kacang-kacangan dan kacang-kacangan lainnya dapat membantu menyeimbangkan gula darah, mengurangi nafsu makan, dan melindungi dari kanker usus besar. Kacang adalah makanan anti-diabetes yang dapat membantu menurunkan berat badan karena dicerna secara lambat, yang memperlambat peningkatan gula darah setelah makan dan membantu mencegah mengidam makanan dengan meningkatkan rasa kenyang. Mengonsumsi kacang-kacangan dan kacang-kacangan lainnya dua kali seminggu telah terbukti mengurangi risiko kanker usus besar. Mengonsumsi kacang-kacangan dan kacang-kacangan lainnya, seperti kacang merah, kacang hitam, buncis, lentil, dan kacang polong, juga memberikan perlindungan yang signifikan terhadap kanker lainnya.
jamur
Makan jamur secara teratur dikaitkan dengan penurunan risiko kanker payudara. Jamur putih dan Portobello sangat bermanfaat melawan kanker payudara karena mengandung aromatase inhibitor atau senyawa yang menghambat produksi estrogen. Jamur telah terbukti memiliki efek anti-inflamasi serta memberikan peningkatan aktivitas sel kekebalan, pencegahan kerusakan DNA, memperlambat pertumbuhan sel kanker, dan penghambatan angiogenesis. Jamur harus selalu dimasak karena jamur mentah memiliki bahan kimia yang berpotensi karsinogenik yang dikenal sebagai agaritine yang berkurang secara signifikan dengan memasak.
Bawang dan Bawang Putih
Bawang dan bawang putih memberikan manfaat sistem kardiovaskular dan kekebalan tubuh serta memberikan efek anti-diabetes dan anti-kanker. Ini juga dikaitkan dengan risiko kanker lambung dan prostat yang lebih rendah. Bawang dan bawang putih dikenal karena senyawa organosulfurnya yang membantu mencegah perkembangan kanker dengan cara mendetoksifikasi karsinogen, mengurangi pertumbuhan sel kanker, dan memblokir angiogenesis. Bawang dan bawang putih juga memiliki konsentrasi tinggi antioksidan flavonoid yang mempromosikan kesehatan, yang memiliki efek anti-inflamasi yang dapat membantu menyediakan pencegahan kanker.
tomat
Tomat kaya akan berbagai nutrisi, seperti likopen, vitamin C dan E, beta-karoten, dan antioksidan flavonol. Lycopene dapat membantu melindungi dari kanker prostat, kerusakan kulit akibat sinar UV, dan? penyakit kardiovaskular. Likopen lebih baik diserap saat tomat dimasak. Satu cangkir saus tomat memiliki sekitar 10 kali jumlah likopen dibandingkan dengan secangkir tomat mentah yang dipotong-potong. Ingatlah juga bahwa karotenoid, seperti likopen, paling baik diserap jika disertai dengan lemak sehat, jadi nikmati tomat Anda dalam salad dengan kacang-kacangan atau saus berbahan dasar kacang untuk mendapatkan manfaat nutrisi tambahan.
Makanan yang kita makan dapat berpotensi bermanfaat atau berbahaya bagi kesehatan kita. Nutrisi yang buruk dapat menyebabkan berbagai masalah kesehatan, termasuk obesitas, penyakit kardiovaskular, dan diabetes tipe 2. Sementara itu, nutrisi yang tepat dapat membuat Anda merasa berenergi, mengurangi risiko masalah kesehatan, serta membantu menjaga dan mengatur berat badan yang sehat. Jika Anda ingin mempromosikan umur panjang, Anda harus mengisi tubuh Anda dengan makanan yang baik. Makanan yang baik juga dapat membantu mengurangi peradangan yang terkait dengan berbagai masalah kesehatan, termasuk nyeri sendi dan radang sendi. Para profesional perawatan kesehatan, seperti ahli tulang, dapat menawarkan saran diet dan gaya hidup untuk membantu meningkatkan kesehatan dan kesejahteraan. Pada artikel berikut, kami akan mencantumkan beberapa makanan enak yang pada akhirnya dapat membantu mempromosikan umur panjang. - Dr. Alex Jimenez DC, CCST Insight
Jus Bit Zesty
Porsi: 1 Waktu memasak: 5-10 menit
1 buah jeruk bali, kupas, iris
1 buah apel, cuci bersih lalu iris-iris
1 buah bit utuh, dan daunnya jika ada, cuci bersih dan iris
1 inchi ruas jahe, dibilas, kupas dan cincang
Jus semua bahan dalam juicer berkualitas tinggi. Terbaik dilayani segera.
Hanya satu wortel memberi Anda semua asupan vitamin A harian Anda
Ya, makan hanya satu wortel 80g (2 oz) rebus memberi Anda cukup beta karoten bagi tubuh Anda untuk menghasilkan 1,480 mikrogram (mcg) vitamin A (diperlukan untuk pembaruan sel kulit). Itu lebih dari asupan harian vitamin A yang direkomendasikan di Amerika Serikat, yaitu sekitar 900mcg. Yang terbaik adalah makan wortel yang dimasak, karena ini melembutkan dinding sel sehingga lebih banyak beta karoten diserap. Menambahkan makanan yang lebih sehat ke dalam diet Anda adalah cara yang bagus untuk meningkatkan kesehatan Anda secara keseluruhan.
Cakupan informasi kami terbatas pada chiropraktik, muskuloskeletal, obat-obatan fisik, kesehatan, dan masalah kesehatan sensitif dan / atau artikel, topik, dan diskusi kedokteran fungsional. Kami menggunakan protokol kesehatan & kebugaran fungsional untuk merawat dan mendukung perawatan cedera atau gangguan pada sistem muskuloskeletal. Pos, topik, subjek, dan wawasan kami mencakup masalah klinis, masalah, dan topik yang terkait dan mendukung secara langsung atau tidak langsung ruang lingkup praktik klinis kami. * Kantor kami telah melakukan upaya yang wajar untuk memberikan kutipan yang mendukung dan telah mengidentifikasi studi penelitian yang relevan atau studi yang mendukung posting kami. Kami juga menyediakan salinan studi penelitian pendukung untuk dewan dan atau publik atas permintaan. Kami memahami bahwa kami mencakup hal-hal yang memerlukan penjelasan tambahan tentang bagaimana hal itu dapat membantu dalam rencana perawatan atau protokol perawatan tertentu; Oleh karena itu, untuk membahas lebih lanjut pokok bahasan di atas, silakan bertanya kepada Dr. Alex Jimenez atau hubungi kami di 915-850-0900. Penyedia Berlisensi di Texas * & New Mexico *
Dikuratori oleh Dr. Alex Jimenez DC, CCST
Referensi:
Joel Fuhrman, MD. 10 Makanan Terbaik yang Dapat Anda Makan untuk Hidup Lebih Lama dan Tetap Sehat. Kesehatan Yang Sangat Baik, 6 Juni 2020, www.verywellhealth.com/best-foods-for-longevity-4005852.
Dowden, Angela. Kopi Adalah Buah dan Fakta Makanan Sejati Luar Biasa Lainnya. Gaya Hidup MSN, 4 Juni 2020, www.msn.com/en-us/foodanddrink/did-you-know/coffee-is-a-fruit-and-other-unbelievably-true-food-facts/ss-BB152Q5q?li=BBnb7Kz&ocid =mailsignout#image=24.
Jika Anda mengalami salah satu dari situasi ini, maka peptida kolagen Anda mungkin rendah.
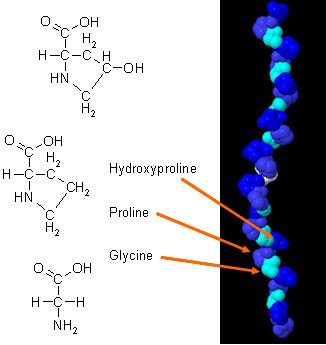
Ada menjadi studi baru tentang bagaimana kolagen dapat meningkatkan komposisi tubuh ketika dikombinasikan dengan latihan harian. Kolagen dalam tubuh memiliki komposisi asam amino unik yang memainkan peran penting dalam anatomi tubuh. Protein kolagen adalah sumber terkonsentrasi glisin, prolin, dan hidroksiprolin, dan ketika sedang dibandingkan dengan semua protein diet lainnya, itu membuat kolagen pilihan praktis yang potensial sebagai protein struktural.
In studi 2015, para peneliti telah menunjukkan bagaimana suplemen kolagen yang efisien dapat meningkatkan komposisi tubuh pada pria aktif. Hasilnya menunjukkan bagaimana setiap individu pria berpartisipasi dalam latihan beban setidaknya tiga kali seminggu dan harus melengkapi dengan setidaknya 15 gram kolagen peptida untuk mencapai kesehatan maksimum. Penilaian yang diberikan oleh tes ini adalah tes kekuatan, analisis bioimpedance (BIA), dan biopsi otot. Tes-tes ini memastikan bahwa individu pria berkinerja baik setelah mengonsumsi suplemen kolagen, dan hasilnya menunjukkan bagaimana massa tubuh mereka mengalami peningkatan massa tubuh bebas lemak. Studi lain menunjukkan bagaimana suplementasi protein kolagen ketika dikombinasikan dengan pelatihan resistensi yang dapat meningkatkan massa otot dan kekuatan otot dengan orang tua serta orang-orang dengan sarcopenia.
Properti Menguntungkan Dengan Kolagen
Ada banyak khasiat yang bermanfaat bahwa suplemen kolagen dapat menyediakan bagi tubuh ketika dikonsumsi. Ada kolagen dan gelatin terhidrolisis dan dapat membantu memperbaiki struktur kulit seseorang. Meskipun tidak ada banyak penelitian tentang suplemen kolagen, ada janji yang sangat baik untuk area pada tubuh. Mereka:
Massa otot: Suplemen kolagen, bila dikombinasikan dengan latihan kekuatan, dapat meningkatkan massa otot dan kekuatan dalam tubuh.
Radang sendi: Suplemen kolagen dapat membantu penderita osteoartritis. Penelitian menunjukkan bahwa ketika orang osteoartritis mengonsumsi suplemen kolagen, mereka menemukan penurunan besar dalam rasa sakit yang mereka alami.
Elastisitas kulit: Di studi 2014, dinyatakan bahwa wanita yang mengonsumsi suplemen kolagen dan telah menunjukkan peningkatan elastisitas kulit. Kolagen juga dapat digunakan dalam perawatan topikal untuk membantu memperbaiki penampilan kulit seseorang dengan meminimalkan garis-garis halus dan kerutan.
Tidak hanya suplemen kolagen yang memberikan khasiat yang bermanfaat bagi area spesifik pada tubuh, tetapi ada empat jenis kolagen utama dan apa peran mereka dalam tubuh manusia serta fungsinya:
Ketik 1: Kolagen tipe 1 memperhitungkan 90% kolagen tubuh dan terdiri dari serat padat yang menyediakan struktur pada kulit, tulang, jaringan ikat, dan gigi yang ada di dalam tubuh.
Ketik 2: Kolagen tipe 2 terdiri dari serat-serat longgar yang ditemukan di tulang rawan elastis, yang membantu melindungi sendi di tubuh.
Ketik 3: Kolagen tipe 3 membantu mendukung struktur otot, organ, dan arteri yang memastikan bahwa tubuh berfungsi dengan benar.
Ketik 4: Kolagen tipe 4 ditemukan di lapisan kulit semua orang dan membantu penyaringan di dalam tubuh.
Karena keempat jenis kolagen ini ada di dalam tubuh, sangat penting untuk mengetahui bahwa kolagen secara alami dapat menurun seiring waktu karena tubuh akan menghasilkan kualitas kolagen yang lebih rendah. Salah satu tanda yang terlihat dari penurunan kolagen adalah ketika kulit pada tubuh manusia menjadi kurang kencang dan kenyal serta melemahkan tulang rawan akibat penuaan.
Faktor-Faktor Yang Dapat Merusak Kolagen
Meskipun kolagen dapat berkurang secara alami seiring bertambahnya usia, banyak faktor yang dapat menghancurkan kolagen yang berbahaya bagi kulit. Faktor-faktor berbahaya dapat meliputi:
Gula dan Karbohidrat: Gula dan karbohidrat olahan dapat mengganggu dengan kemampuan kolagen untuk memperbaiki dirinya sendiri pada kulit. Sehingga dengan meminimalisir konsumsi gula dan karbohidrat dalam tubuh dapat mengurangi efek disfungsi pembuluh darah, ginjal, dan jaringan kulit.
Paparan Matahari: Meskipun mendapatkan cukup sinar matahari dapat membantu seseorang menikmati hari, bagaimanapun, terkena sinar matahari untuk waktu yang lama dapat menyebabkan kerusakan ke kulit dan menghancurkan peptida kolagen. Efek paparan sinar matahari yang berlebihan dapat menyebabkan kulit memotret usia dan menghasilkan stres oksidatif dalam tubuh.
Merokok: Ketika seseorang merokok, itu bisa mengurangi produksi kolagen di dalam tubuh, menyebabkan tubuh memiliki keriput dini, dan jika tubuh terluka, proses penyembuhan akan lebih lambat dan dapat menyebabkan penyakit di tubuh.
Penyakit autoimun: Beberapa penyakit autoimun juga dapat merusak produksi kolagen seperti lupus.
Kesimpulan
Kolagen sangat penting bagi tubuh karena membantu kulit menjadi lembut dan kencang. Secara alami, ini akan berkurang seiring bertambahnya usia seseorang, jadi mengonsumsi suplemen kolagen dapat memastikan bahwa tubuh dapat berfungsi dengan benar. Ketika faktor-faktor berbahaya mempengaruhi tubuh, mereka dapat menghentikan atau bahkan merusak produksi kolagen dan mempercepat proses keriput dini dari pembentukan, membuat seseorang terlihat lebih tua daripada mereka. Beberapa produk dapat membantu aktivitas seluler tubuh dengan memberikan stabilitas yang lebih baik, ketersediaan hayati, dan kenyamanan pencernaan.
Ruang lingkup informasi kami terbatas pada masalah chiropraktik, muskuloskeletal, dan saraf atau artikel fungsional kedokteran, topik, dan diskusi. Kami menggunakan protokol kesehatan fungsional untuk mengobati cedera atau gangguan pada sistem muskuloskeletal. Kantor kami telah melakukan upaya yang wajar untuk memberikan kutipan yang mendukung dan telah mengidentifikasi studi penelitian yang relevan atau studi yang mendukung posting kami. Kami juga membuat salinan studi penelitian pendukung tersedia untuk dewan dan atau publik atas permintaan. Untuk lebih lanjut membahas masalah di atas, jangan ragu untuk bertanya kepada Dr. Alex Jimenez atau hubungi kami di 915-850-0900.
Referensi:
Bosch, Ricardo, dkk. Mekanisme Fotoaging dan Fotokarsinogenesis Kulit, dan Strategi Fotoprotektif dengan Fitokimia. Antioksidan (Basel, Swiss), MDPI, 26 Maret 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4665475/.
Danby, F William. Nutrisi dan Penuaan Kulit: Gula dan Glikasi Klinik di Dermatologi, Perpustakaan Kedokteran Nasional AS, 2010, www.ncbi.nlm.nih.gov/pubmed/20620757.
Jennings, Kerri-Ann. Collagen - What Is It dan What Is It Good For? Healthline, 9 September 2016, www.healthline.com/nutrition/collagen.
Jurgelewicz, Michael. Penelitian Baru Mendemonstrasikan Manfaat Peptida Kolagen untuk Meningkatkan Komposisi Tubuh yang Dikombinasikan dengan Latihan. Desain untuk Kesehatan, 31 Mei 2019, blog.designsforhealth.com/node/1031.
Knuutinen, A, dkk. Merokok Mempengaruhi Sintesis Kolagen dan Perputaran Matriks Ekstraseluler pada Kulit Manusia. British Journal of Dermatology, Perpustakaan Kedokteran Nasional AS, Apr. 2002, www.ncbi.nlm.nih.gov/pubmed/11966688.
Proksch, E, dkk. Suplementasi oral dari Peptida Kolagen Tertentu Memiliki Efek Menguntungkan pada Fisiologi Kulit Manusia: Studi Double-Blind, Placebo-Controlled. Farmakologi dan Fisiologi Kulit, Perpustakaan Kedokteran Nasional AS, 2014, www.ncbi.nlm.nih.gov/pubmed/23949208.
Schauss, Alexander G, dkk. Efek dari Ekstrak Tulang Rawan Bagian Dalam Ayam yang Dihidrolisis dengan Berat Molekul Rendah, Kolagen BioCell, pada Peningkatan Gejala Terkait Osteoartritis: Uji Coba Acak, Buta Ganda, Terkontrol Plasebo. Jurnal Kimia Pertanian dan Pangan, Perpustakaan Kedokteran Nasional AS, 25 Apr. 2012, www.ncbi.nlm.nih.gov/pubmed/22486722.
Zdzieblik, Denise, dkk. Suplementasi Peptida Kolagen dalam Kombinasi dengan Latihan Ketahanan Memperbaiki Komposisi Tubuh dan Meningkatkan Kekuatan Otot pada Pria Lansia Sarkopen: Percobaan Terkontrol Secara Acak. The British Journal of Nutrition, Cambridge University Press, 28 Oktober 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4594048/.
Dengan memberi tahu individu-individu tentang bagaimana Universitas Nasional Ilmu Kesehatan memberikan pengetahuan untuk generasi masa depan, Universitas menawarkan berbagai macam profesi medis untuk kedokteran fungsional.
Seperti Anda telah didiagnosis menderita Penyakit Celiac, Irritable Bowel Syndrome, Diverticulosis / Diverticulitis, atau Leaky Gut Syndrome?
Bersendawa berlebihan, bersendawa, atau kembung?
Distensi abnormal setelah probiotik atau suplemen alami tertentu?
Kecurigaan malabsorpsi gizi?
Apakah masalah pencernaan mereda dengan relaksasi?
Jika Anda mengalami salah satu dari situasi ini, maka Anda mungkin mengalami masalah usus dan mungkin harus mencoba Program 4R.
Sensitivitas makanan, rheumatoid arthritis, dan kecemasan telah dikaitkan dengan gangguan permeabilitas gastrointestinal. Berbagai kondisi ini dapat terjadi dari banyak faktor yang dapat berdampak pada saluran pencernaan. Jika tidak diobati, hal ini berpotensi sebagai hasil dari disfungsi penghalang permeabilitas usus, menyebabkan peradangan, dan kondisi kesehatan yang parah sehingga usus dapat berkembang. Program 4R digunakan untuk memulihkan usus yang sehat di dalam tubuh dan melibatkan empat langkah. Mereka adalah: menghapus, mengganti, menginstal ulang, dan memperbaiki.
Permeabilitas Usus
Permeabilitas usus membantu melindungi tubuh dan memastikan bahwa bakteri berbahaya tidak memasuki usus. Ini melindungi tubuh dari faktor lingkungan potensial yang bisa berbahaya dan masuk melalui saluran pencernaan. Ini bisa berupa toksin, mikroorganisme patogen, dan antigen lain yang dapat merusak saluran pencernaan yang menyebabkan masalah. Lapisan usus terdiri dari lapisan sel epitel yang dipisahkan oleh persimpangan ketat. Dalam usus yang sehat, persimpangan ketat mengatur permeabilitas usus dengan secara selektif membiarkan zat masuk dan melakukan perjalanan melintasi penghalang usus dan mencegah faktor-faktor berbahaya dari diserap.
Faktor lingkungan tertentu dapat merusak persimpangan ketat, dan hasilnya adalah itu dapat meningkatkan permeabilitas usus, yang menyebabkan hiperpermeabilitas usus atau usus bocor dalam tubuh. Faktor yang berkontribusi dapat meningkatkan permeabilitas usus seperti jumlah lemak jenuh dan alkohol yang berlebihan, kekurangan nutrisi, stres kronis, dan penyakit menular.
Dengan permeabilitas usus yang meningkat di dalam usus, itu dapat memungkinkan antigen untuk menyeberangi mukosa usus dan memasuki aliran darah yang menyebabkan respon imun dan peradangan pada tubuh. Ada beberapa kondisi pencernaan tertentu yang berhubungan dengan hiperpermeabilitas usus dan jika tidak diobati dapat memicu kondisi autoimun tertentu yang dapat menyebabkan kerusakan pada tubuh.
Program 4Rs
4Rs adalah program yang disarankan para profesional kesehatan untuk digunakan pasien mereka ketika mereka sedang menangani masalah pencernaan yang mengganggu dan membantu mendukung penyembuhan usus.
Menghapus Masalah
Langkah pertama dalam program 4R adalah untuk menghilangkan patogen berbahaya dan pemicu peradangan yang terkait dengan peningkatan permeabilitas usus. Pemicu seperti stres dan konsumsi alkohol kronis dapat banyak merusak tubuh seseorang. Jadi penargetan faktor-faktor berbahaya dari tubuh ini adalah mengobatinya dengan obat-obatan, antibiotik, suplemen, dan penghilangan makanan radang dari makanan yang disarankan, termasuk:
- Alkohol
- Perekat
- Aditif makanan
- Pati
- Asam lemak tertentu
- Makanan tertentu yang sensitif bagi seseorang
Mengganti Nutrisi
Langkah kedua dari program 4R adalah mengganti nutrisi yang menyebabkan masalah usus melalui peradangan. Nutrisi tertentu dapat membantu mengurangi peradangan di usus sambil memastikan bahwa saluran pencernaan didukung. Ada beberapa makanan antiinflamasi yang bergizi. Ini termasuk:
- Makanan tinggi serat
- Omega-3s
- Minyak zaitun
- Jamur
- Herbal anti-inflamasi
Ada suplemen tertentu yang dapat digunakan untuk mendukung fungsi pencernaan dengan membantu dan menyerap nutrisi untuk mempromosikan usus yang sehat. Apa yang dilakukan oleh enzim pencernaan adalah membantu dalam memecah lemak, protein, dan karbohidrat dalam usus. Ini akan membantu memberi manfaat pada individu yang mengalami gangguan saluran pencernaan, intoleransi makanan, atau memiliki penyakit celiac. Suplemen seperti suplemen asam empedu dapat membantu membantu penyerapan nutrisi dengan menggabungkan lemak bersama. Penelitian telah menyatakan bahwa asam empedu telah digunakan untuk mengobati hati, kantong empedu, dan saluran empedu sambil mencegah pembentukan batu empedu setelah operasi bariatrik.
Reinokulasi Usus
Langkah ketiga adalah program 4rs untuk menginokulasi ulang mikroba usus dengan bakteri menguntungkan untuk mempromosikan fungsi usus yang sehat. Penelitian telah ditunjukkan bahwa suplemen probiotik telah digunakan untuk memperbaiki usus dengan memulihkan bakteri menguntungkan. Dengan suplemen ini, mereka memberikan peningkatan usus dengan mengeluarkan zat anti-inflamasi ke dalam tubuh, membantu mendukung sistem kekebalan tubuh, mengubah komposisi mikroba tubuh, dan mengurangi permeabilitas usus dalam sistem usus.
Sejak probiotik ditemukan dalam makanan yang difermentasi dan dianggap sementara karena mereka tidak bertahan lama di saluran pencernaan dan bermanfaat. Anehnya, mereka masih berdampak pada kesehatan manusia karena mempengaruhi usus dengan memproduksi vitamin dan senyawa anti-mikroba, sehingga memberikan keragaman dan fungsi usus.
Memperbaiki Usus
Langkah terakhir program 4R adalah memperbaiki usus. Langkah ini melibatkan perbaikan lapisan usus usus dengan nutrisi spesifik dan herbal. Tumbuhan dan suplemen ini dapat membantu mengurangi permeabilitas usus dan peradangan dalam tubuh. Beberapa herbal dan suplemen ini meliputi:
- Lidah buaya
- Chios permen karet
- DGL (liclyice Deglycyrrhizinated)
- Akar Marshmallow
- L-glutamin
- Omega-3s
Polifenol
- Vitamin D
- Seng
Kesimpulan
Karena banyak faktor yang dapat mempengaruhi sistem pencernaan dengan cara yang berbahaya dan dapat menjadi kontributor beberapa kondisi kesehatan. Tujuan utama dari program 4R adalah untuk meminimalkan faktor-faktor yang merusak usus dan mengurangi peradangan dan meningkatkan permeabilitas usus. Ketika pasien diperkenalkan dengan faktor-faktor bermanfaat yang diberikan 4R, itu dapat mengarah pada usus yang sehat dan sembuh. Beberapa produk hadir untuk membantu mendukung sistem pencernaan dengan mendukung usus, meningkatkan metabolisme gula, dan menargetkan asam amino yang dimaksudkan untuk mendukung usus.
Ruang lingkup informasi kami terbatas pada masalah chiropraktik, muskuloskeletal, dan saraf atau artikel fungsional kedokteran, topik, dan diskusi. Kami menggunakan protokol kesehatan fungsional untuk mengobati cedera atau gangguan pada sistem muskuloskeletal. Kantor kami telah melakukan upaya yang wajar untuk memberikan kutipan yang mendukung dan telah mengidentifikasi studi penelitian yang relevan atau studi yang mendukung posting kami. Kami juga membuat salinan studi penelitian pendukung tersedia untuk dewan dan atau publik atas permintaan. Untuk lebih lanjut membahas masalah di atas, jangan ragu untuk bertanya kepada Dr. Alex Jimenez atau hubungi kami di 915-850-0900.
Referensi:
De Santis, Stefania, dkk. Tombol Nutrisi untuk Modulasi Penghalang Usus. Frontiers dalam Imunologi, Frontiers Media SA, 7 Desember 2015, www.ncbi.nlm.nih.gov/pmc/articles/PMC4670985/.
Ianiro, Gianluca, dkk. Suplementasi Enzim Pencernaan pada Penyakit Gastrointestinal Metabolisme Obat Saat Ini, Penerbit Ilmu Bentham, 2016, www.ncbi.nlm.nih.gov/pmc/articles/PMC4923703/.
Mu, Qinghui, dkk. Leaky Gut Sebagai Sinyal Bahaya untuk Penyakit Autoimun. Frontiers, Frontiers, 5 Mei 2017, www.frontiersin.org/articles/10.3389/fimmu.2017.00598/full.
Rezac, Shannon, dkk. Makanan Fermentasi sebagai Sumber Makanan Organisme Hidup Perbatasan dalam Mikrobiologi, Frontiers Media SA, 24 Agustus 2018, www.ncbi.nlm.nih.gov/pmc/articles/PMC6117398/.
Sander, Guy R., dkk. Gangguan Cepat Fungsi Penghalang Usus oleh Gliadin Melibatkan Perubahan Ekspresi Protein Junctional Apikal. FEBS Tekan, John Wiley & Sons, Ltd, 8 Agustus 2005, febs.onlinelibrary.wiley.com/doi/full/10.1016/j.febslet.2005.07.066.
Sartor, R Balfour. Manipulasi Terapeutik Mikroflora Enterik dalam Penyakit Radang Usus: Antibiotik, Probiotik, dan Prebiotik. Gastroenterologi, Perpustakaan Kedokteran Nasional AS, Mei 2004, www.ncbi.nlm.nih.gov/pubmed/15168372.
Nyeri kronis adalah masalah kesehatan umum yang mempengaruhi banyak orang di Amerika Serikat. Sementara beberapa kondisi medis, seperti fibromyalgia dan sindrom nyeri myofascial, dapat menyebabkan nyeri kronis, itu juga dapat berkembang karena berbagai masalah kesehatan lainnya. Studi penelitian telah menemukan bahwa peradangan yang meluas adalah penyebab utama nyeri kronis. Peradangan adalah mekanisme pertahanan alami terhadap cedera, penyakit, atau infeksi. Tapi, jika proses inflamasi berlanjut terlalu lama, itu bisa menjadi masalah.
Peradangan menandakan sistem kekebalan tubuh untuk menyembuhkan dan memperbaiki jaringan yang rusak serta untuk melindungi diri terhadap bakteri dan virus. Seperti disebutkan di atas, bagaimanapun, peradangan kronis dapat menyebabkan berbagai masalah kesehatan, termasuk gejala nyeri kronis. Modifikasi gaya hidup sehat dapat membantu mengelola nyeri kronis, tetapi pertama-tama, mari kita pahami penyebab umum nyeri kronis.
Apa itu Inflamasi Akut?
Peradangan akut, misalnya, terjadi setelah cedera atau sesuatu yang sederhana seperti sakit tenggorokan. Ini adalah respons alami dengan efek buruk, yang berarti ini bekerja secara lokal di wilayah tempat masalah kesehatan ditemukan. Tanda-tanda umum dari peradangan akut termasuk pembengkakan, kemerahan, kehangatan, rasa sakit dan kehilangan fungsi, sebagaimana dinyatakan oleh Perpustakaan Kedokteran Nasional. Ketika peradangan akut berkembang, pembuluh darah melebar menyebabkan aliran darah meningkat, dan sel darah putih di daerah yang terluka meningkatkan pemulihan.
Selama peradangan parah, senyawa yang disebut sitokin dilepaskan oleh jaringan yang rusak. Sitokin bertindak sebagai "sinyal darurat" yang membawa sel-sel kekebalan tubuh manusia sendiri, serta hormon dan banyak nutrisi untuk memperbaiki masalah kesehatan. Selain itu, zat seperti hormon, yang dikenal sebagai prostaglandin, menyebabkan pembekuan darah untuk menyembuhkan jaringan yang rusak, dan ini juga dapat memicu demam dan rasa sakit sebagai bagian dari prosedur inflamasi. Saat kerusakan atau cedera pulih, peradangan mereda.
Apa itu Peradangan Kronis?
Tidak seperti peradangan akut, peradangan kronis memiliki efek jangka panjang. Peradangan kronis, juga dikenal sebagai peradangan persisten, menghasilkan peradangan tingkat rendah di seluruh tubuh manusia, seperti yang ditunjukkan oleh peningkatan penanda sistem kekebalan yang terletak di darah dan jaringan sel. Peradangan kronis juga dapat menyebabkan perkembangan berbagai penyakit dan kondisi. Peningkatan tingkat peradangan kadang-kadang dapat memicu bahkan jika tidak ada cedera, penyakit, atau infeksi, yang juga dapat menyebabkan sistem kekebalan tubuh bereaksi.
Akibatnya, sistem kekebalan tubuh manusia bisa mulai menyerang sel, jaringan, atau organ yang sehat. Para peneliti masih mencoba memahami konsekuensi dari peradangan kronis pada tubuh manusia dan mekanisme yang terlibat dalam proses pertahanan alami ini. Misalnya, peradangan kronis telah dikaitkan dengan berbagai masalah kesehatan, seperti penyakit jantung, dan stroke.
Satu teori menunjukkan bahwa ketika peradangan tetap di pembuluh darah, itu dapat mendorong akumulasi plak. Menurut American Heart Association, atau AHA, jika sistem kekebalan mengidentifikasi plak sebagai penyerbu asing, sel-sel darah putih dapat mencoba untuk membendung plak yang ditemukan dalam darah yang mengalir melalui arteri. Ini dapat membuat gumpalan darah yang dapat menghalangi aliran darah ke jantung atau otak, menyebabkannya menjadi tidak stabil dan pecah. Kanker adalah masalah kesehatan lain yang terkait dengan peradangan kronis. Lebih lanjut, menurut National Cancer Institute, kerusakan DNA juga bisa disebabkan oleh peradangan kronis.
Peradangan yang menetap dan tingkat rendah sering tidak memiliki gejala, tetapi para profesional kesehatan dapat memeriksa protein C-reaktif, atau CRP, yang dikenal sebagai asam lipoat, penanda untuk peradangan yang ditemukan dalam darah. Peningkatan kadar CRP dikaitkan dengan peningkatan risiko penyakit kardiovaskular. Peningkatan kadar CRP dapat ditemukan pada gangguan kronis seperti lupus atau rheumatoid arthritis.
Dalam kasus kondisi kronis lainnya, seperti fibromyalgia, sistem saraf bereaksi berlebihan terhadap rangsangan spesifik, namun, peradangan yang menyebabkan gejala nyeri kronis. Secara subyektif, hampir tidak mungkin untuk mengetahui perbedaan antara nyeri kronis yang disebabkan oleh sistem saraf yang terlalu sensitif dan nyeri kronis yang disebabkan oleh peradangan yang meluas. Selain mencari petunjuk dalam aliran darah, nutrisi seseorang, kebiasaan gaya hidup, dan paparan lingkungan, juga dapat meningkatkan peradangan kronis.
Peradangan adalah mekanisme pertahanan alami sistem kekebalan terhadap cedera, penyakit, atau infeksi. Meskipun respons peradangan ini dapat membantu menyembuhkan dan memperbaiki jaringan, peradangan kronis yang meluas dapat menyebabkan berbagai masalah kesehatan, termasuk gejala nyeri kronis. Seimbang nutrisi, termasuk berbagai diet dan puasa, dapat membantu mengurangi peradangan. Puasa, juga dikenal sebagai pembatasan kalori, meningkatkan apoptosis sel dan pemulihan mitokondria. Diet meniru puasa, yang merupakan bagian dari rencana diet umur panjang, adalah program diet yang “mengelabui” tubuh manusia ke dalam keadaan berpuasa untuk merasakan manfaat dari puasa tradisional. Sebelum mengikuti diet apa pun yang dijelaskan dalam artikel ini, pastikan untuk berkonsultasi dengan dokter.
Dr Alex Jimenez DC, CCST Insight
Nutrisi, Makanan, Puasa dan Nyeri Kronis
Diet anti-inflamasi terutama terdiri dari makan buah-buahan dan sayuran segar, ikan, dan lemak. Rencana diet Mediterania, misalnya, adalah diet anti-inflamasi yang mempromosikan makan kacang dalam jumlah sedang, mengonsumsi sedikit daging, dan minum anggur. Bagian makanan anti-inflamasi, seperti asam lemak omega-3, melindungi tubuh manusia terhadapnya dabijaksana disebabkan oleh peradangan.
Diet anti-inflamasi juga melibatkan menjauhi makanan yang bisa memicu peradangan. Ini sangat ideal untuk mengurangi jumlah makanan yang Anda makan yang tinggi lemak trans dan jenuh, seperti daging. Selain itu, diet antiinflamasi membatasi konsumsi karbohidrat olahan dan makanan, seperti roti dan nasi. Ini juga mempromosikan mengurangi pemanfaatan margarin dan minyak yang dikemas dengan asam lemak omega-6, seperti bunga matahari, safflower dan minyak jagung.
Puasa, atau pembatasan kalori, telah lama diketahui mengurangi stres oksidatif dan memperlambat mekanisme penuaan pada berbagai organisme. Efek puasa melibatkan kematian sel yang diprogram, atau apoptosis, transkripsi, efisiensi energi seluler, biogenesis mitokondria, mekanisme antioksidan, dan ritme sirkadian. Puasa juga berkontribusi terhadap autophagy mitokondria, yang dikenal sebagai mitofag, di mana gen dalam mitokondria dirangsang untuk menjalani apoptosis, yang mendorong pemulihan mitokondria.
Puasa intermiten dapat membantu Anda melawan peradangan, meningkatkan pencernaan, dan meningkatkan umur panjang Anda. Tubuh manusia dirancang untuk dapat bertahan dalam waktu lama tanpa makanan. Studi penelitian telah menunjukkan bahwa puasa intermiten dapat memiliki perubahan positif pada keseluruhan komposisi mikrobiota usus Anda. Apalagi puasa intermiten dapat menurunkan resistensi insulin sekaligus meningkatkan respon sistem imun. Akhirnya, puasa intermiten dapat meningkatkan produksi zat, yang dikenal sebagai? -Hydroxybutyrate, yang memblokir sebagian dari sistem kekebalan yang terlibat dalam penyakit inflamasi serta secara substansial mengurangi produksi penanda inflamasi, seperti sitokin dan protein C-reaktif. , atau CRP, yang disebutkan sebelumnya di atas.
Rencana Diet Panjang Umur, yang disajikan dalam buku oleh Dr. Valter Longo, menghilangkan konsumsi makanan olahan yang dapat menyebabkan peradangan, meningkatkan kesejahteraan dan umur panjang. Program diet unik ini, tidak seperti kebanyakan diet tradisional, tidak mempromosikan penurunan berat badan. Meskipun Anda mungkin mengalami penurunan berat badan, penekanan program diet unik ini adalah makan lebih sehat. Longevity Diet Plan telah terbukti membantu mengaktifkan pembaruan berbasis sel induk, mengurangi lemak perut, dan mencegah keropos tulang dan otot yang berkaitan dengan usia, serta membangun ketahanan terhadap pengembangan penyakit kardiovaskular, penyakit Alzheimer, diabetes, dan kanker.
Diet menirukan puasa, atau PMK, memungkinkan Anda untuk merasakan manfaat puasa tradisional tanpa mengurangi makanan tubuh Anda. Perbedaan utama dari PMK adalah bahwa alih-alih menghilangkan semua makanan selama beberapa hari atau bahkan berminggu-minggu, Anda hanya membatasi asupan kalori selama lima hari dalam sebulan. PMK dapat dilakukan sebulan sekali untuk membantu meningkatkan kesehatan dan kesejahteraan secara keseluruhan.
Sementara siapa pun dapat mengikuti PMK sendiri, namun ProLon diet meniru puasa menawarkan program makan 5-hari yang telah dikemas dan diberi label tersendiri untuk setiap hari, yang menyajikan makanan yang Anda butuhkan untuk PMK dalam jumlah dan kombinasi yang tepat. Program makan terdiri dari makanan nabati yang siap makan atau mudah disiapkan, termasuk bar, sup, camilan, suplemen, konsentrat minuman, dan teh. Sebelum memulai ProLon diet meniru puasa, program makan 5 hari, atau modifikasi gaya hidup yang dijelaskan di atas, pastikan untuk berbicara dengan profesional kesehatan untuk mengetahui perawatan nyeri kronis mana yang tepat untuk Anda.
Ruang lingkup informasi kami terbatas pada chiropraktik, masalah kesehatan tulang belakang, dan artikel, topik, dan diskusi kedokteran fungsional. Untuk membahas lebih lanjut masalah pokok di atas, silakan bertanya kepada Dr. Alex Jimenez atau hubungi kami di 915-850-0900 .
Diundangkan oleh Dr. Alex Jimenez
Diskusi Topik Tambahan: Sakit Punggung Akut
Nyeri punggung adalah salah satu penyebab kecacatan yang paling umum dan hari-hari yang terlewatkan di dunia kerja. Atribut nyeri punggung menjadi alasan paling umum kedua untuk kunjungan dokter, kalah jumlah hanya oleh infeksi saluran pernapasan atas. Sekitar 80 persen dari populasi akan mengalami sakit punggung setidaknya satu kali sepanjang hidup mereka. Tulang belakang Anda adalah struktur kompleks yang terdiri dari tulang, sendi, ligamen, dan otot, di antara jaringan lunak lainnya. Cedera dan / atau kondisi yang memburuk, seperti cakram hernia, akhirnya dapat menyebabkan gejala nyeri punggung. Cedera olahraga atau cedera kecelakaan mobil sering menjadi penyebab paling sering dari nyeri punggung, namun terkadang gerakan yang paling sederhana dapat memiliki hasil yang menyakitkan. Untungnya, pilihan pengobatan alternatif, seperti perawatan chiropractic, dapat membantu meringankan nyeri punggung melalui penggunaan penyesuaian tulang belakang dan manipulasi manual, yang pada akhirnya meningkatkan pereda nyeri.
XYMOGEN Formula Profesional Eksklusif tersedia melalui profesional perawatan kesehatan berlisensi tertentu. Penjualan dan diskon formula XYMOGEN di internet sangat dilarang.
Dengan bangga, Dr. Alexander Jimenez membuat formula XYMOGEN hanya tersedia untuk pasien di bawah perawatan kami.
Silakan hubungi kantor kami agar kami dapat memberikan konsultasi dokter untuk akses segera.
Jika Anda seorang pasien Klinik Cedera Medis & Chiropractic, Anda dapat menanyakan tentang XYMOGEN dengan menelepon 915-850-0900.
Untuk kenyamanan Anda dan ulasan tentang XYMOGEN produk silakan tinjau tautan berikut. *XYMOGEN-Katalog-Unduh
Mengikuti diet tertentu untuk mempertahankan nutrisi yang tepat terkadang dapat membuat makan menjadi stres. Modifikasi gaya hidup alami adalah kunci untuk mengubah kebiasaan makan Anda dan ini dapat membantu Anda menjalani hidup yang lebih lama, lebih sehat. The Longevity Diet Plan, yang dibuat oleh Dr. Valter Longo, adalah pilihan pedoman makan praktis yang berfokus pada perubahan pola makan Anda untuk mencapai kesehatan dan kesejahteraan secara keseluruhan.
Aturan Rencana Diet Panjang Umur
Dengan hanya mengikuti tips nutrisi di bawah ini, Anda dapat merombak rencana diet Anda saat ini dan mulai makan lebih sehat tanpa semua tekanan dari diet tradisional. Rencana Diet Panjang Umur menghilangkan konsumsi makanan olahan yang dapat menyebabkan berbagai masalah kesehatan dan meningkatkan konsumsi nutrisi yang mendukung umur panjang. Program diet unik ini membagikan hasil sekitar 25 tahun studi penelitian semua pada solusi sederhana yang dapat membantu orang mengalami kesejahteraan secara keseluruhan melalui nutrisi yang tepat.
Namun, tidak seperti kebanyakan diet tradisional, Longevity Diet Plan tidak mendorong penurunan berat badan. Meskipun Anda mungkin mengalami penurunan berat badan, penekanan dari program diet unik ini adalah makan yang lebih sehat. Rencana Diet Panjang Umur telah terbukti membantu Anda mengaktifkan pembaruan berbasis sel induk, menurunkan berat badan dan mengurangi lemak perut, mencegah kehilangan tulang dan otot terkait usia, membangun ketahanan terhadap penyakit kardiovaskular, penyakit Alzheimer, diabetes, dan kanker, juga sebagai memperpanjang umur panjang. Di bawah ini, kami akan merangkum 8 tip nutrisi paling umum dari Rencana Diet Panjang Umur yang pada akhirnya dapat membantu membuat hidup Anda lebih panjang dan lebih sehat.
The Longevity Diet Plan adalah program diet unik yang dirancang oleh Dr. Valter Longo untuk mempromosikan kesehatan, kebugaran, dan umur panjang secara keseluruhan. Melalui modifikasi gaya hidup sederhana, orang dapat mengubah kebiasaan makan mereka dan memanfaatkan banyak manfaat kesehatan dari program diet ini. Dengan mengikuti diet pescatarian dan mengikuti ProLon Diet Meniru Puasa, di antara tips nutrisi lain yang dijelaskan di bawah ini, orang dapat hidup lebih lama dan lebih sehat. Diet tradisional seringkali sulit dan membuat stres untuk diikuti, namun, Longevity Diet Plan adalah program diet yang praktis dan unik yang cocok untuk banyak orang.
Dr Alex Jimenez DC, CCST Insight
Tips Gizi 8 dari Rencana Diet Panjang Umur
Ikuti Diet Pescatarian
Sebagai bagian dari Longevity Diet Plan, ikuti diet pescatarian, yang hampir 100 persen berbasis tanaman dan ikan. Juga, pastikan untuk membatasi konsumsi ikan hingga dua atau tiga porsi setiap minggu, menghindari ikan dengan kandungan merkuri yang lebih tinggi, seperti tuna, ikan pedang, mackerel, dan halibut. Jika Anda lebih dari 65 dan mulai mengalami pengurangan massa otot, kekuatan, dan lemak, tambahkan lebih banyak ikan ke dalam makanan Anda bersama makanan hewani lainnya, termasuk telur dan keju tertentu, seperti feta atau pecorino, dan yogurt yang terbuat dari kambing susu.
Jangan Makan Terlalu Banyak Protein
Menurut Longevity Diet Plan, kita harus mengonsumsi 0.31 hingga 0.36 gram protein per pon lemak tubuh setiap hari. Jika Anda menimbang 130lbs, Anda harus mengonsumsi sekitar 40 hingga 47 gram protein per hari, atau setara dengan 1.5 filet salmon, 1 cangkir buncis atau 2 1 / 2 gelas lentil, di mana 30 gram harus dikonsumsi dalam satu kali makan. Jika Anda menimbang 200 hingga 220lbs, Anda harus makan sekitar 60 hingga 70 gram protein per hari, atau setara dengan dua fillet salmon, 3 1 / 2 cangkir lentil atau 1 1 / 2 cangkir buncis. Konsumsi protein harus ditingkatkan setelah usia 65. Bagi kebanyakan dari kita, peningkatan 10 ke 20 persen, atau 5 ke 10 gram lebih banyak setiap hari, sudah cukup. Akhirnya, Longevity Diet bebas dari protein hewani seperti daging merah, daging putih, dan unggas, dengan pengecualian protein hewani pada ikan. Sebaliknya, program diet yang unik ini kaya protein nabati seperti kacang-kacangan dan kacang-kacangan untuk mengoptimalkan kesehatan dan kesejahteraan.
Tingkatkan Lemak Baik dan Karbohidrat Kompleks
Sebagai bagian dari Rencana Diet Panjang Umur, Anda harus makan lemak tak jenuh ganda dalam jumlah yang lebih tinggi, seperti yang ditemukan pada salmon, almond, kacang kenari, dan minyak zaitun, sedangkan Anda harus mengonsumsi lemak jenuh, hidrogenasi, dan lemak trans dalam jumlah yang lebih rendah. Demikian juga, sebagai bagian dari Rencana Diet Panjang Umur, Anda juga harus makan karbohidrat kompleks, seperti yang ditemukan dalam roti gandum, kacang-kacangan, dan sayuran. Pastikan untuk membatasi makan pasta, nasi, roti, buah, dan jus buah, yang dapat dikonversi menjadi gula pada saat mereka mencapai usus Anda.
Ambil Suplemen Diet
Tubuh manusia membutuhkan protein, asam lemak esensial seperti omega-3 dan omega-6, vitamin, mineral, dan bahkan gula agar berfungsi dengan benar. Setiap kali asupan nutrisi tertentu menjadi terlalu rendah, perbaikan, penggantian, dan metode pertahanan tubuh manusia dapat melambat atau berhenti, memungkinkan jamur, bakteri, dan virus menyebabkan kerusakan yang dapat menyebabkan berbagai masalah kesehatan. Konsumsi suplemen vitamin dan mineral, terutama untuk omega-3, seperti yang direkomendasikan oleh profesional kesehatan Anda.
Makan Berbagai Makanan dari A Andancestry
Untuk mengambil semua nutrisi yang diperlukan yang Anda butuhkan, Anda harus makan berbagai macam makanan, tetapi yang terbaik adalah memilih makanan yang umum di meja orang tua, kakek-nenek, dan kakek-nenek buyut Anda. Sebagai contoh, di banyak negara Eropa utara di mana susu telah dikonsumsi secara umum, intoleransi laktosa relatif jarang, sedangkan intoleransi laktosa cukup umum di negara-negara Eropa selatan dan Asia, di mana susu secara historis bukan bagian dari makanan konvensional orang dewasa. Jika seseorang keturunan Jepang yang tinggal di Amerika Serikat tiba-tiba memutuskan untuk mulai minum susu, yang mungkin jarang disajikan di meja makan kakek-nenek mereka, mereka mungkin akan mulai merasa sakit. Masalah yang paling umum dalam kasus ini adalah intoleransi atau autoimunitas, seperti respon terhadap makanan kaya gluten seperti roti dan pasta yang terlihat pada orang dengan penyakit celiac. Meskipun bukti lebih lanjut diperlukan, ada kemungkinan bahwa intoleransi makanan dapat dikaitkan dengan banyak gangguan autoimun, termasuk diabetes, kolitis, dan penyakit Crohn.
Makan Dua Makanan Sehari dan Camilan
Menurut Rencana Diet Panjang Umur, sangat ideal untuk makan sarapan dan satu kali makan utama ditambah camilan rendah kalori dan rendah gula bergizi setiap hari. Sementara bagi sebagian orang mungkin disarankan untuk makan tiga kali dan satu kali camilan setiap hari. Banyak pedoman nutrisi yang menganjurkan agar kita makan lima hingga enam kali makan setiap hari. Ketika orang disarankan untuk sering makan, seringkali menjadi sulit bagi mereka untuk mengatur asupan kalori mereka. Selama dua puluh tahun terakhir, sekitar 70 persen populasi di Amerika Serikat dianggap kelebihan berat badan atau obesitas. Jauh lebih sulit untuk makan berlebihan pada Rencana Diet Panjang Umur jika Anda hanya makan dua setengah kali sehari. Butuh porsi besar kacang-kacangan, sayuran, dan ikan untuk mencapai jumlah yang akan menyebabkan penambahan berat badan. Makanan yang bergizi tinggi, ditambah jumlah makanannya, mengirimkan sinyal ke perut dan otak Anda bahwa Anda sudah cukup makan. Sistem makanan utama yang satu ini terkadang harus dipecah menjadi dua kali makan untuk menghindari masalah pencernaan. Orang dewasa dan orang tua yang rentan terhadap penurunan berat badan harus makan tiga kali sehari. Untuk orang yang mencoba menurunkan berat badan serta orang yang kelebihan berat badan atau obesitas, saran nutrisi terbaik adalah sarapan setiap hari; makan malam atau makan siang, tetapi tidak keduanya, dan gantikan makan yang terlewat dengan satu camilan yang mengandung kurang dari 100 kalori dan tidak lebih dari 3 hingga 5 g gula. Makanan yang Anda lewatkan bergantung pada gaya hidup Anda, namun, tidak disarankan untuk melewatkan sarapan karena masalah kesehatan yang merugikan. Manfaat melewatkan makan siang adalah lebih banyak waktu dan energi luang. Tapi, ada kekurangan makan malam besar, terutama bagi orang yang menderita refluks asam atau masalah tidur. Namun, kelemahan melewatkan makan malam adalah hal itu dapat menghilangkan jamuan makan sosial pada hari mereka.
Makanlah dalam Jendela 12-Jam Setiap Hari
Kebiasaan makan umum lainnya yang diadopsi oleh banyak centenarian adalah makan dengan waktu terbatas atau membatasi semua makanan dan makanan ringan dalam jendela berjam-jam 12 setiap hari. Efisiensi metode ini ditunjukkan dalam studi penelitian pada manusia dan hewan. Secara umum, Anda akan makan sarapan di 8 pagi dan kemudian makan malam dengan 8 sore. Jendela makan yang lebih singkat sepuluh jam atau kurang bisa menjadi lebih baik untuk menurunkan berat badan, tetapi itu jauh lebih sulit untuk dipertahankan dan mungkin meningkatkan risiko mengembangkan efek samping, seperti batu empedu dan bahkan berpotensi meningkatkan kemungkinan mengembangkan penyakit kardiovaskular. Anda sebaiknya tidak makan tiga hingga empat jam sebelum tidur.
Ikuti Diet Meniru Puasa ProLon®
Orang sehat di bawah usia 65 harus mengikuti ProLon Fasting Mimicking Diet, program makan 5 hari setidaknya dua kali setiap tahun. FMD adalah salah satu prinsip utama yang dipromosikan oleh Longevity Diet Plan. Diet meniru puasa menawarkan manfaat kesehatan yang sama dari puasa tanpa benar-benar puasa. Dengan mengonsumsi 800 hingga 1,100 kalori dalam jumlah yang tepat dan kombinasi makanan yang telah dikemas secara individual dan diberi label untuk setiap hari, Anda dapat "mengelabui" tubuh manusia ke keadaan puasa. Melalui berbagai studi penelitian, Dr. Valter Longo menemukan bahwa dengan menghilangkan tubuh makanan dengan cara ini, sel-sel kita mulai memecah dan meregenerasi jaringan internal kita, melalui proses yang dikenal sebagai autophagy, membunuh dan mengganti, atau regenerasi, sel yang rusak. Selain itu, puasa dapat membalikkan berbagai masalah kesehatan, menghancurkan sel-sel kanker dan secara signifikan mengurangi kemungkinan mengembangkan penyakit Alzheimer.
Dengan Rencana Diet Panjang Umur yang disajikan dalam buku oleh Dr. Valter Longo, Anda akan makan lebih baik, merasa lebih baik dan, meskipun itu tidak dirancang sebagai rencana penurunan berat badan, Anda bahkan dapat menurunkan berat badan beberapa kilogram. Anda tidak perlu mempertimbangkan aturan makanan yang rumit dan membuat pilihan sulit dengan program diet unik ini. Setelah Anda memahami modifikasi gaya hidup ini, Anda juga akan dapat meningkatkan kesehatan dan kebugaran Anda secara keseluruhan Tujuan umur panjang. Grafik ruang lingkup informasi kami terbatas pada chiropraktik, masalah kesehatan tulang belakang, dan topik pengobatan fungsional. Untuk membahas lebih lanjut masalah ini, silakan bertanya kepada Dr. Alex Jimenez atau hubungi kami di 915-850-0900 .
Diundangkan oleh Dr. Alex Jimenez
Diskusi Topik Tambahan: Sakit Punggung Akut
Nyeri punggung adalah salah satu penyebab kecacatan yang paling umum dan hari-hari yang terlewatkan di dunia kerja. Atribut nyeri punggung menjadi alasan paling umum kedua untuk kunjungan dokter, kalah jumlah hanya oleh infeksi saluran pernapasan atas. Sekitar 80 persen dari populasi akan mengalami sakit punggung setidaknya satu kali sepanjang hidup mereka. Tulang belakang Anda adalah struktur kompleks yang terdiri dari tulang, sendi, ligamen, dan otot, di antara jaringan lunak lainnya. Cedera dan / atau kondisi yang memburuk, seperti cakram hernia, akhirnya dapat menyebabkan gejala nyeri punggung. Cedera olahraga atau cedera kecelakaan mobil sering menjadi penyebab paling sering dari nyeri punggung, namun terkadang gerakan yang paling sederhana dapat memiliki hasil yang menyakitkan. Untungnya, pilihan pengobatan alternatif, seperti perawatan chiropractic, dapat membantu meringankan nyeri punggung melalui penggunaan penyesuaian tulang belakang dan manipulasi manual, yang pada akhirnya meningkatkan pereda nyeri.
XYMOGEN Formula Profesional Eksklusif tersedia melalui profesional perawatan kesehatan berlisensi tertentu. Penjualan dan diskon formula XYMOGEN di internet sangat dilarang.
Dengan bangga, Dr. Alexander Jimenez membuat formula XYMOGEN hanya tersedia untuk pasien di bawah perawatan kami.
Silakan hubungi kantor kami agar kami dapat memberikan konsultasi dokter untuk akses segera.
Jika Anda seorang pasien Klinik Cedera Medis & Chiropractic, Anda dapat menanyakan tentang XYMOGEN dengan menelepon 915-850-0900.
Untuk kenyamanan Anda dan ulasan tentang XYMOGEN produk silakan tinjau tautan berikut. *XYMOGEN-Katalog-Unduh
Alat Temukan Praktisi IFM adalah jaringan rujukan terbesar dalam Pengobatan Fungsional, dibuat untuk membantu pasien menemukan praktisi Pengobatan Fungsional di mana pun di dunia. Praktisi Bersertifikat IFM terdaftar pertama dalam hasil pencarian, mengingat pendidikan ekstensif mereka dalam Kedokteran Fungsional