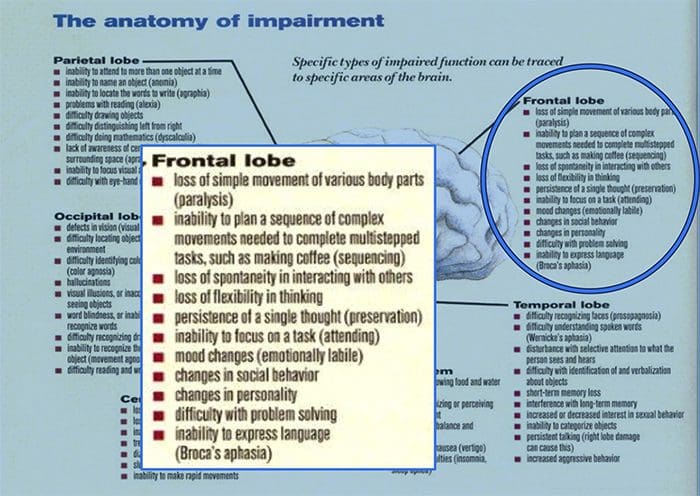
Memotong Saraf Anda Mengubah Otak Anda | El Paso, TX.
Setelah transeksi saraf tepi ekstremitas atas dan operasi perbaikan, beberapa pasien mendapatkan kembali fungsi sensorimotor yang baik sementara yang lain tidak. Memahami mekanisme perifer dan sentral yang berkontribusi pada pemulihan dapat memfasilitasi pengembangan intervensi terapeutik baru. Plastisitas setelah transeksi saraf tepi telah dibuktikan di seluruh neuroaksis pada model hewan dari cedera saraf. Namun, perubahan otak yang terjadi setelah transeksi saraf tepi dan perbaikan bedah pada manusia belum diperiksa. Lebih lanjut, sejauh mana regenerasi saraf perifer mempengaruhi perubahan fungsional dan struktural otak belum dikarakterisasi. Oleh karena itu, kami bertanya apakah perubahan fungsional disertai dengan perubahan struktural materi abu-abu dan / atau putih dan apakah perubahan ini terkait dengan pemulihan sensorik? Untuk mengatasi masalah utama ini kami (i) menilai regenerasi saraf perifer; (ii) mengukur aktivasi otak pencitraan resonansi magnetik fungsional (sinyal tergantung tingkat oksigen darah; BOLD) sebagai respons terhadap stimulus vibrotaktil; (iii) memeriksa plastisitas struktur otak materi abu-abu dan putih; dan (iv) tindakan pemulihan sensorik berkorelasi dengan perubahan materi abu-abu pada pasien transeksi saraf tepi dan perbaikan bedah. Dibandingkan dengan saraf kontralesi sehat setiap pasien, saraf yang ditranseksi telah merusak konduksi saraf 1.5 tahun setelah transeksi dan perbaikan, konduksi dengan amplitudo yang menurun dan latensi yang meningkat. Dibandingkan dengan kontrol yang sehat, transeksi saraf tepi dan pasien perbaikan bedah telah mengubah aktivitas sinyal tergantung pada tingkat oksigen darah di korteks somatosensori primer dan sekunder kontralesi, dan dalam satu set area otak yang dikenal sebagai `` jaringan positif tugas ''. Selain itu, pengurangan materi abu-abu diidentifikasi di beberapa area otak, termasuk korteks somatosensori primer dan sekunder kontralesi, di area yang sama di mana pengurangan sinyal yang bergantung pada tingkat oksigen darah diidentifikasi. Selain itu, penipisan materi abu-abu pada girus pasca-pusat berkorelasi negatif dengan pengukuran pemulihan sensorik (deteksi mekanis dan getaran) yang menunjukkan hubungan yang jelas antara fungsi dan struktur. Akhirnya, kami mengidentifikasi pengurangan anisotropi pecahan materi putih di insula kanan di wilayah yang juga menunjukkan berkurangnya materi abu-abu. Hasil ini memberikan wawasan tentang plastisitas otak dan hubungan struktur-fungsi-perilaku setelah cedera saraf dan memiliki implikasi terapeutik yang penting.
Kata kunci: ketebalan kortikal; fMRI; pencitraan tensor difusi; keliatan; cedera saraf perifer
Singkatan: BA = area Brodmann; BOLD = tingkat oksigen darah tergantung; fMRI = pencitraan resonansi magnetik fungsional;
PNIr = transeksi saraf perifer dan perbaikan bedah; S1 = korteks somatosensori primer; S2 = korteks somatosensori sekunder
Pengantar
Setelah transeksi saraf perifer ekstremitas atas dan perbaikan bedah (PNIr), 25% pasien tidak kembali bekerja 1.5 tahun setelah operasi (Jaquet et al., 2001). Selain itu, ?57% pasien dengan cedera saraf berusia antara 16-35 tahun (McAllister et al., 1996); dengan demikian, kecacatan seumur hidup dan kesulitan ekonomi dapat menyertai transeksi saraf ekstremitas atas. Memahami konsekuensi sentral dan perifer dari cedera saraf perifer dapat memfasilitasi pengembangan strategi terapi baru dan program intervensi.
Tidak diketahui bagaimana otak merespon PNIr pada manusia. Namun, penelitian pada hewan telah menetapkan bahwa plastisitas dalam korteks somatosensori dimulai segera setelah transeksi saraf perifer, dan bahwa 1 tahun setelah transeksi saraf lengkap dan perbaikan bedah, peta kortikal mengandung tambalan, representasi yang tidak berkesinambungan dari saraf yang ditransplantasikan dan berdekatan (Wall et al. ., 1986). Mekanisme yang memfasilitasi plastisitas fungsional diperkirakan mencakup pembukaan segera proyeksi yang sudah ada dari tingkat kortikal dan subkortikal yang berdekatan, dan jangka panjang tunas akson pada berbagai tingkat neuroaxis, termasuk korteks somatosensori primer (S1) (Florence dan Kaas , 1995; Hickmott dan Steen, 2005).
Studi pencitraan otak manusia telah menguatkan temuan dari model hewan dengan identifikasi peta aktivasi MRI fungsional akibat cedera tulang belakang, amputasi, transfer to-thumb, dan pada pasien dengan sindrom terowongan karelel (Lotze et al., 2001; Manduch et al., 2002; Jurkiewicz et al., 2006; Napadow et al., 2006). Selanjutnya, penelitian struktural MRI baru-baru ini memvisualisasikan perubahan materi abu-abu dan putih berikut cedera traumatis dan dalam berbagai kondisi patologis, termasuk amputasi ekstremitas dan nyeri kronis (Apkarian dkk., 2004; Draganski dkk., 2006; Davis et al., 2008; Geha et al., 2008; May, 2008). Perubahan materi abu-abu dianggap terkait dengan perubahan ukuran sel, atrofi dan / atau hilangnya neuron atau glia, sedangkan perubahan materi putih dipengaruhi oleh degenerasi aksonal dan hilangnya myelin (Beaulieu, 2002; May, 2008).
Pendekatan yang kuat untuk menggambarkan mekanisme patologi dan plastisitas adalah dengan menggabungkan teknik pencitraan fungsional berwarna abu-abu dan putih fungsional dan struktural. Kami sebelumnya melaporkan bahwa pasien dengan PNIr ekstremitas atas lengkap mempertahankan defisit somatosensori mendalam yang bertahan selama 41.5 setelah operasi (Taylor et al., 2008a). Berdasarkan temuan ini, kami beralasan pasien ini akan menunjukkan perubahan otak fungsional dan struktural di daerah otak somatosensorik kunci. Oleh karena itu, dalam penelitian ini, kami berhipotesis bahwa pasien PNIr akan memiliki: (i) mengurangi respon tergantung kadar oksigen darah (BOLD) terhadap stimulasi vibrasi dari wilayah saraf yang ditransmisikan, di wilayah S1 yang mewakili ekstremitas atas yang cedera dan di sekunder. korteks somatosensori (S2); (ii) pengurangan yang sesuai dalam ketebalan kortikal di wilayah ini dari S1 kontralesi dan S2; (Iii) korelasi antara perubahan ketebalan kortikal dan pengukuran psikofisik fungsi somatosensori (getaran dan ambang deteksi sentuhan); dan (iv) mengurangi fraksional anisotropi (ukuran integritas zat putih) dalam materi putih yang dimasukkan ke dalam / keluar dari area kortikal somatosensori ini.
metode
Subjek
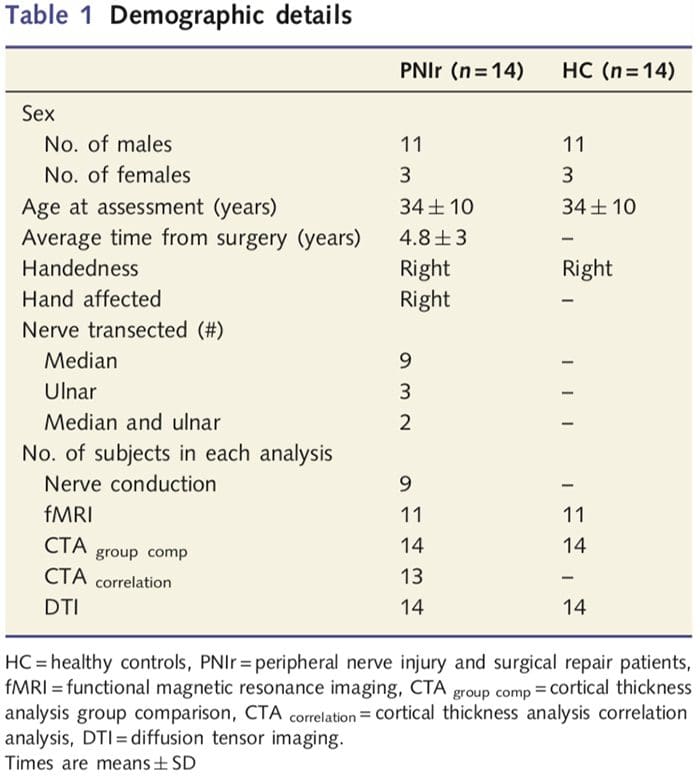
Kami merekrut 27 pasien dengan transeksi lengkap saraf median dan/atau ulnaris diikuti dengan perbaikan bedah dari ahli bedah plastik yang berafiliasi dengan University of Toronto Hand Program antara Juni 2006 dan Mei 2008. Dari kohort yang lebih besar ini, 14 pasien bebas rasa sakit (tiga perempuan , 11 laki-laki; 34 ? 10 tahun) dengan transeksi lengkap nervus medianus dan/atau ulnaris kanan dimasukkan dalam penelitian [untuk menghindari pembaur terkait dengan adanya nyeri dan pasien lateralitas dengan nyeri (n=6) dan lesi sisi kiri (n=7) dikeluarkan dari analisis ini]. Semua pasien menjalani perbaikan saraf bedah mikro setidaknya 1.5 tahun sebelum pendaftaran studi (waktu pemulihan bervariasi 1.5-8 tahun). Selain itu, kami merekrut 14 kontrol sehat dengan usia dan jenis kelamin yang sesuai (3 perempuan, 11 laki-laki; 34 ? 10 tahun). Semua subjek memberikan persetujuan tertulis yang diinformasikan untuk prosedur yang disetujui oleh Dewan Etika Penelitian Jaringan Kesehatan Universitas. Semua subjek menggunakan tangan kanan (ditentukan dengan menggunakan inventaris tangan kanan Edinburgh: Oldfield, 1971) dan tidak memiliki riwayat cedera neurologis atau nyeri kronis (baik sebelum atau setelah transeksi saraf). Lihat Tabel 1 untuk detail demografis.
Desain studi
Semua subjek berpartisipasi dalam sesi pencitraan yang meliputi: (i) pencitraan resonansi magnetik fungsional (fMRI) sebagai respons terhadap rangsangan vibrotactile yang diterapkan pada jari telunjuk kanan (dalam wilayah saraf median); (ii) pemindaian anatomi resolusi tinggi dari seluruh otak, diperoleh untuk registrasi citra dan untuk analisis materi abu-abu kortikal; dan (iii) dua difusi pemindaian tensor pencitraan untuk penilaian integritas zat putih. Sebelum pencitraan, subjek diinstruksikan dalam desain dasar eksperimen dan diingatkan untuk tetap diam selama durasi pemindaian.
Subjek bebas untuk menarik diri dari studi setiap saat. Selain itu, penilaian sensorik dan motorik dilakukan untuk semua subjek (Taylor et al., 2008a). Karena ambang deteksi sentuhan dan getaran berkorelasi dengan ketebalan kortikal, deskripsi metode ini termasuk di bawah ini (tindakan psikofisik lainnya akan dilaporkan di tempat lain).
Ambang Vibrasi
Ambang deteksi getaran ditentukan menggunakan Bio-Thesiometer genggam (Bio-Medical Instrument Company, USA). Perangkat ini memiliki probe 12-mm yang ditempatkan pada phalanx distal jari telunjuk kanan (D2). Ambang batas ditentukan dengan menggunakan metode batas: amplitudo (tegangan) secara bertahap meningkat sampai subjek menunjukkan bahwa mereka merasakan stimulus. Ambang getaran diperoleh tiga kali dan nilai rata-rata dihitung. Selama pengujian getaran ambang, subjek diperintahkan untuk menutup mata mereka dan menyandarkan punggung tangan mereka pada bantalan yang mendukung.
Ambang Deteksi Mekanis
Ambang batas deteksi mekanis ditentukan menggunakan satu set standar filamen von Frey (OptiHair2 Marstock Nervtest, Jerman) yang berisi 12 filamen kalibrasi dengan jarak logaritmik yang mengirimkan gaya dari 0.25-512 mN. Diameter permukaan kontak dari semua 12 filamen adalah ~ 0.4mm. Uji coba dilakukan dengan mata tertutup dan tangan diletakkan di atas bantal yang empuk. Probe diterapkan dalam rangkaian menaik dan subjek diminta untuk membuat respons setiap kali mereka merasakan probe menyentuh ujung jari kanan D2. Proses ini diulangi tiga kali. Gaya untuk filamen yang terdeteksi setidaknya dalam dua dari tiga percobaan dilaporkan sebagai ambang deteksi mekanis subjek.
Pemeriksaan Konduksi Saraf
Pasien berpartisipasi dalam studi konduksi saraf motorik dan sensorik bilateral di klinik elektromiografi Rumah Sakit Toronto Western (EMG). Untuk konduksi saraf motorik, elektroda perangsang ditempatkan di pergelangan tangan dan siku (secara terpisah) dan elektroda perekam ditempatkan di atas penculik pollicis brevis, untuk pemeriksaan saraf median, atau digiti minimi penculik untuk pemeriksaan saraf ulnaris. Untuk pengujian saraf sensorik, elektroda perekam ditempatkan di pergelangan tangan dan elektroda perangsang ditempatkan di digit D2, D3 dan D5. Seorang ahli saraf senior yang berpengalaman dari Klinik EMG Rumah Sakit Barat Toronto (Dr Peter Ashby) meninjau semua penilaian klinis untuk menentukan saraf mana yang menunjukkan respons normal / abnormal. Karena ukuran amplitudo dan latensi diketahui bervariasi secara substansial antara subjek (karena faktor-faktor seperti kepadatan persarafan, kedalaman saraf dan ketebalan kulit subjek individu) (Kimura, 2001) setiap saraf pasien yang tidak ditranseksi berfungsi sebagai kontrol mereka sendiri untuk perbandingan dengan nilai-nilai dari sisi yang ditranseksi. Pada pasien dengan respons konduksi saraf yang dapat dideteksi, uji-t berpasangan dilakukan untuk menilai perbedaan dalam ukuran latensi atau amplitudo antara setiap saraf pasien yang ditranseksi dan kontralesi yang tidak ditranseksi.
Parameter Imaging
Data pencitraan otak diperoleh menggunakan sistem 3T GE MRI yang dilengkapi dengan kumparan kepala array bertahap delapan saluran. Subyek ditempatkan terlentang di atas meja MRI dan kepala masing-masing subjek diberi bantalan untuk mengurangi gerakan. Data fMRI seluruh otak diperoleh menggunakan pencitraan echo planar (28 irisan aksial, bidang pandang (FOV) = 20 x 20 cm, matriks 64 x 64, voxel 3.125 x 3.125 x 4mm, waktu gema (TE) = 30 ms, pengulangan waktu (TR) = 2000 ms). Waktu pemindaian adalah 5 menit dan 8 detik (154 frame). Selama pemindaian, stimulus vibrotactile 12 Hz yang tidak menyakitkan diterapkan ke falang distal D2 kanan menggunakan diafragma balon yang digerakkan oleh udara terkompresi (Perangkat diproduksi oleh Dr Christo Pantev; www.biomag.uni-muenster.de). Stimuli diberikan dalam blok 10-an yang disisipkan dengan 20-an istirahat, dengan total 10 blok stimulasi dan 10 blok istirahat. Data 8 detik (4 TR) pertama yang diperoleh dari setiap proses dibuang untuk memungkinkan ekuilibrasi sinyal fMRI. Subjek diinstruksikan untuk menutup mata selama pemindaian dan fokus pada rangsangan. Pemindaian anatomis resolusi tinggi tiga dimensi (3D) seluruh otak (124 irisan sagital, 24 x 24 cm FOV, 256 x 256 matriks, 1.5 x 0.94 x 0.94 mm voxel) diperoleh dengan urutan gema gradien manja 1D berbobot T3 (satu sinyal rata-rata, sudut balik = 20? , TE ?5 ms). Selain itu, dua pemindaian pencitraan tensor difusi (38 irisan aksial, FOV 24 x 24 cm, matriks 128 x 128, voxel 1.875 x 1.875 x 3 mm) diperoleh di sepanjang 23 arah dengan nilai b 1000smm�2. Setiap run juga berisi dua volume tanpa bobot difusi.
Analisis fMRI
Data dianalisis menggunakan Brainvoyager QX v1.8 (Brain Innovaton, Maastricht, Belanda). Pra-pemrosesan termasuk: koreksi gerak 3D, koreksi slice scan-time, penghapusan tren linear, penyaringan high-pass (lima siklus per run), dan smoothing spasial dengan lebar penuh 6mm pada setengah maksimum (FWHM) kernel Gaussian. set data fMRI diinterpolasi ke 3 x 3 x 3 mm voxels, terdaftar pada gambar anatomi resolusi tinggi, dan dinormalkan ke ruang Talairach standar (Talairach dan Tournoux, 1988). Voxel dilaporkan sebagai 1 x 1 x 1 mm. Data dianalisis menggunakan model linear umum; model ini diperoleh dengan mengubah fungsi boxcar dari kursus stimulasi taktil dengan fungsi respon hemodinamik standar. Untuk mengidentifikasi antara perbedaan kelompok dalam pola aktivasi, analisis efek tetap dilakukan dengan kontras: (i) kontrol yang sehat: stimulasi 4 istirahat; (ii) PNIr: stimulasi 4 istirahat; dan (iii) kontrol sehat 4 PNIr. Peta aktivasi di-threshold pada nilai P50.05 yang telah dikoreksi (berasal dari PoxNUMX yang tidak dikoreksi dan 50.0001mm120 voxels yang bersebelahan seperti yang dilaporkan sebelumnya: Taylor dan Davis, 3); ini juga divalidasi dengan menjalankan Simulasi Monte Carlo dengan aplikasi AlphaSim diimplementasikan dalam perangkat lunak Analisis Fungsional Neuroimage (AFNI). Analisis ini hanya melibatkan pasien 2009 yang melanjutkan transeksi nervus median kanan (n = 11) atau median kanan dan saraf ulnar (n = 9) (yaitu tiga pasien dengan transeksi nervus ulnaris kanan murni tidak dimasukkan dalam analisis ini. ).
Analisis Ketebalan Kortikal
Analisis ketebalan kortikal dilakukan dengan menggunakan Freesurfer (http: // surfer.nmr.mgh.harvard.edu); metode telah diuraikan secara rinci di tempat lain (Dale et al., 1999; Fischl et al., 1999a, b; Fischl dan Dale 2000). Secara singkat, set data anatomi T1-weighted resolusi tinggi telah didaftarkan ke atlas Talairach (Talairach dan Tournoux, 1988). Ini diikuti dengan normalisasi intensitas, pengupasan tengkorak dan pemisahan belahan. Selanjutnya, batas materi putih / abu-abu (disebut permukaan putih) dan abu-abu / CSF (disebut permukaan pial) diidentifikasi dan disegmentasi. Jarak antara permukaan putih dan pial kemudian dihitung di setiap titik di setiap belahan otak. Untuk mengidentifikasi perbedaan kelompok antara 14 pasien dan 14 kontrol yang sesuai usia / jenis kelamin, analisis model linier umum dilakukan pada setiap titik di otak. Karena topografi kortikal individu secara inheren heterogen, kernel penghalusan spasial 5mm FWHM diterapkan sebelum analisis statistik. Data ditampilkan pada P50.05 yang dikoreksi (berasal dari simpul bersebelahan P50.0075 dan 102 yang tidak dikoreksi); ini dihitung dengan menjalankan simulasi Monte Carlo dengan AlphaSim. Sebuah simpul merepresentasikan sebuah titik pada lembaran dua dimensi, dan, dalam studi ini, jarak antara dua simpul adalah 0.80mm2.
Karena pasien menunjukkan defisit yang signifikan dalam fungsi somatosensori dalam wilayah saraf yang ditranseksi, kami berhipotesis bahwa pengukuran fungsi somatosensori (deteksi getaran dan sentuhan) akan berkorelasi dengan ketebalan kortikal pada gyrus pasca-sentral kontralesi (korteks somatosensori primer dan sekunder). Oleh karena itu, kami melakukan analisis korelasi pada kelompok pasien antara: (i) ketebalan kortikal dan ambang deteksi getaran; dan (ii) ketebalan kortikal dan ambang deteksi sentuhan. Satu pasien tidak menyelesaikan penilaian psikofisik; oleh karena itu, analisis ini melibatkan 13 pasien PNIr. Selain itu, untuk mengetahui apakah ada hubungan antara ketebalan kortikal dan waktu pemulihan dilakukan juga analisis korelasi antara kedua pengukuran tersebut. Analisis korelasi ini dibatasi pada girus pasca-pusat kontralesi dengan memasukkan sungkup (diambil dari atlas bawaan Freesurfer) dalam model linier umum. Simulasi Monte Carlo dilakukan yang dibatasi pada jumlah simpul dalam gyrus pasca-sentral kontralesi; gambar ditampilkan dengan P50.05 yang dikoreksi (berasal dari simpul bersebelahan P50.0075 dan 68 yang tidak dikoreksi).
Analisis Pencitraan Difusi Tensor
Pemrosesan citra tensor difusi dilakukan dengan DTiStudio (www.MriStudio.org) dan FSLv.4.0 (www.fmrib.ox.ac.uk/fsl/). Gambar pertama kali diselaraskan dengan alat Registrasi Gambar Otomatis yang diimplementasikan di DTiStudio, menggunakan gambar B0 pertama di seri pertama yang diperoleh sebagai template. Proses ini mengoreksi gerakan subjek dan distorsi arus pusar. Semua gambar kemudian diperiksa secara visual untuk menilai kualitas gambar dan penyelarasan dari pencitraan tensor difusi yang terpisah berjalan. Jika artefak terdeteksi, potongan telah dihapus sebelum menghitung rata-rata dari dua pencitraan tensor difusi yang terpisah. Peta FA individu dihitung menggunakan alat DTIFIT yang diterapkan di FSL. Analisis statistik Voxel-bijaksana dilakukan untuk mengidentifikasi perbedaan kelompok dalam anisotropi fraksional rata-rata menggunakan Statistik Spasial Berbasis Tract; untuk penjelasan lengkap tentang metode ini lihat Smith et al. (2006). Secara singkat, gambar didaftarkan secara non-linier ke gambar target (MNI152), gambar rata-rata kemudian dibuat dari semua dataset dan gambar ini kemudian diencerkan untuk mewakili semua bidang yang umum untuk semua subjek. Nilai anisotropi pecahan tertinggi masing-masing subjek kemudian diproyeksikan ke kerangka dengan mencari di materi putih tegak lurus setiap titik pada kerangka materi putih. Analisis statistik seluruh otak berdasarkan voxel kemudian dilakukan antara kelompok (14PNIr dan 14 kontrol sehat) dan gambar seluruh otak dikoreksi pada P50.05. Selain itu, analisis wilayah minat dilakukan di materi putih traktat berdekatan dengan S1 kontralateral, thalamus dan insula anterior dan posterior bilateral. Daerah-daerah ini dipilih karena mereka sebelumnya telah terlibat dalam aspek somatosensasi dan karena mereka sesuai dengan daerah yang diidentifikasi dalam analisis fMRI dan analisis ketebalan kortikal (CTA). Daerah yang tertarik digambar pada white matter skeleton sebagai berikut: (i) Wilayah S1 kontralateral yang menarik berasal medial di persimpangan antara white matter skeleton korona-radiata dan bagian skeleton makan ke gyrus pasca sentral; mengakhiri pada akhir saluran dalam irisan yang diberikan.
Dalam arah z wilayah yang diinginkan diperluas dari z = 49 menjadi 57; saluran materi putih memasok wilayah tangan. (ii) Daerah talamus kontralateral terbatas pada jejak materi putih yang mengelilingi inti talamus posterior dan medial (inti yang terlibat dalam fungsi somatosensori), membentang dari z = 1 sampai 4. (iii) Daerah insular yang menarik digambar secara bilateral di dalamnya materi putih berdekatan dengan insula anterior dan posterior berdasarkan kriteria yang sebelumnya diterbitkan oleh lab kami (Taylor et al., 2008b). Wilayah minat diperpanjang dari z = 2 menjadi 8. Nilai anisotropi pecahan diekstraksi dari masing-masing wilayah minat ini dan analisis varians multivariat (MANOVA) dilakukan dengan menggunakan Paket Statistik untuk Ilmu Sosial v13.0 (SPSS Inc, Chicago), yang menyertakan nilai anisotropi pecahan untuk keenam wilayah yang diminati.
Hasil
Tabel 1 memberikan rincian demografis untuk peserta studi. Semua 14 pasien mengalami transeksi lengkap dari median kanan dan/atau saraf ulnaris diikuti oleh perbaikan bedah mikro setidaknya 1.5 tahun sebelum pendaftaran studi. Waktu dari operasi hingga pengujian berkisar antara 1.5 hingga 8 tahun dengan rata-rata (?SD) 4.8 ? 3 tahun. Pasien dan kontrol tidak (34 ? 10 tahun kedua kelompok; t = 0.04; P = 0.97).
Psikofisika
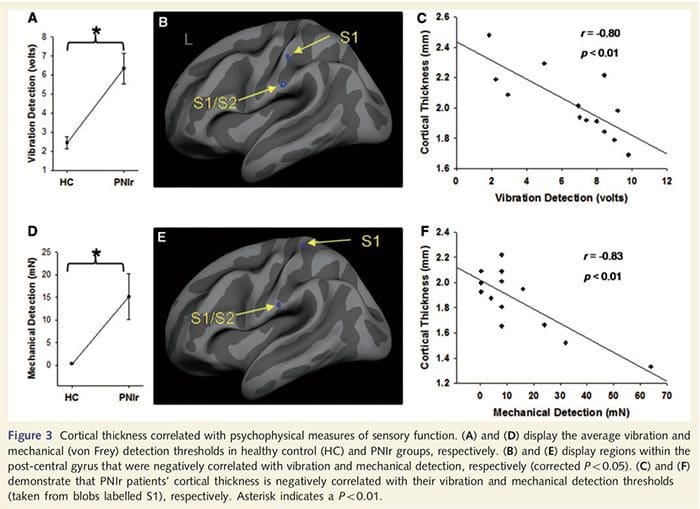
Ambang getaran dihitung dari ketiga pengukuran sejak analisis pengukuran varians satu arah yang diulang (ANOVA) menunjukkan tidak ada perbedaan signifikan antara ketiga percobaan [F (25, 1) = 0.227, P = 0.64]. Getaran dan ambang deteksi mekanis secara signifikan terganggu pada pasien PNIr dibandingkan dengan kontrol yang sehat (getaran: t = 4.77, P50.001, Gambar. 3A; mekanik: t = 3.10, P = 0.005, Gambar. 3D).
Pemeriksaan Konduksi Saraf
Ukuran amplitudo dan latensi yang diperoleh dari setiap saraf kontralesi pasien diklasifikasikan sebagai normal oleh ahli saraf berpengalaman di Klinik EMG Rumah Sakit Toronto Western. Sembilan dari 14 pasien menyelesaikan pengujian konduksi saraf. Tabel 2 menampilkan rata-rata peningkatan / penurunan latensi dan data amplitudo untuk konduksi saraf sensorik dari pergelangan tangan ke otot abduktor pollicis brevis (median) atau penculik digiti minimi (ulnar) dan untuk konduksi sensorik dari pergelangan tangan ke D2 (median) dan D5 ( ulnaris) dibandingkan dengan setiap pasien yang tidak cedera saraf kontralesi. Dari sembilan, tujuh pasien mengalami transeksi termasuk saraf median. Dari ketujuh pasien tersebut, satu pasien tidak memiliki respons yang terdeteksi selama pengujian motorik dan pasien lain tidak memiliki respons yang terdeteksi selama pengujian sensorik.
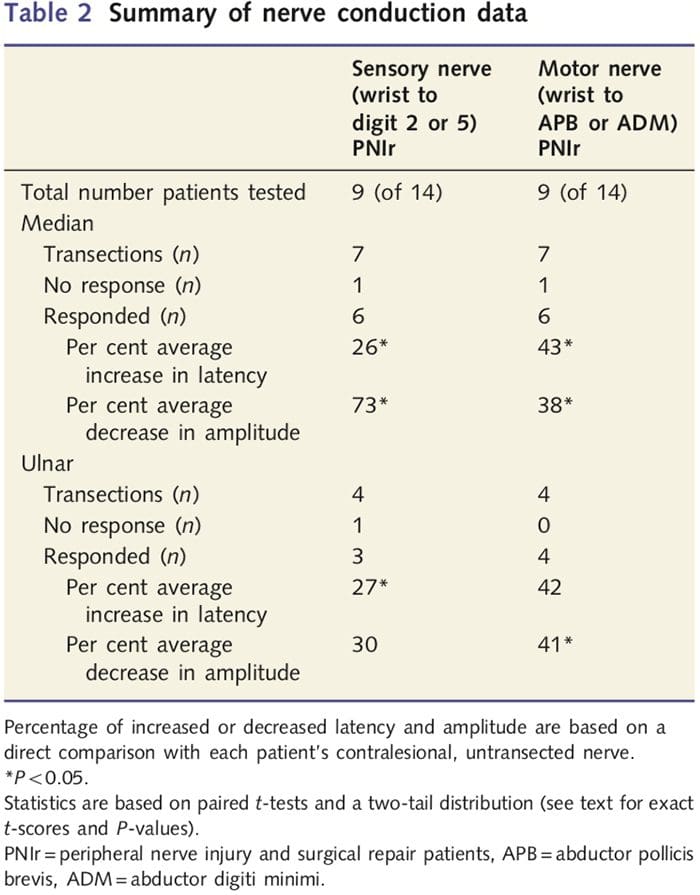 Pada enam pasien dengan respon yang dapat dideteksi, latensi konduksi motorik meningkat sebesar 43% (t = 6.2; P = 0.002) dan amplitudo menurun sebesar 38% (t = 2.6; P = 0.045) ketika setiap saraf pasien yang ditranseksi dibandingkan dengan sisi mereka yang tidak terluka. Konduksi sensorik pada saraf median juga menunjukkan peningkatan latensi sebesar 26% (t = 3.9; P = 0.011) dan penurunan amplitudo sebesar 73% (t = 8.0; P = 0.000) dibandingkan dengan saraf kontralesi normal. Pada empat pasien dengan transeksi saraf ulnaris, satu pasien tidak memiliki respons yang terdeteksi selama pengujian saraf sensorik. Pada pasien dengan respon, latensi motorik saraf ulnaris tidak meningkat secara signifikan (t = 2.8; P = 0.070); Namun, amplitudo secara signifikan
Pada enam pasien dengan respon yang dapat dideteksi, latensi konduksi motorik meningkat sebesar 43% (t = 6.2; P = 0.002) dan amplitudo menurun sebesar 38% (t = 2.6; P = 0.045) ketika setiap saraf pasien yang ditranseksi dibandingkan dengan sisi mereka yang tidak terluka. Konduksi sensorik pada saraf median juga menunjukkan peningkatan latensi sebesar 26% (t = 3.9; P = 0.011) dan penurunan amplitudo sebesar 73% (t = 8.0; P = 0.000) dibandingkan dengan saraf kontralesi normal. Pada empat pasien dengan transeksi saraf ulnaris, satu pasien tidak memiliki respons yang terdeteksi selama pengujian saraf sensorik. Pada pasien dengan respon, latensi motorik saraf ulnaris tidak meningkat secara signifikan (t = 2.8; P = 0.070); Namun, amplitudo secara signifikan
menurun 41% (t = 5.9; P = 0.010). Pengujian sensorik saraf ulnaris menunjukkan peningkatan latensi 27% (t = 4.3; P = 0.049) tetapi tidak ada peningkatan amplitudo yang signifikan (t =? 3.5; P = 0.072).
Plastisitas Fungsional Dalam Korteks Somatosensori Primer
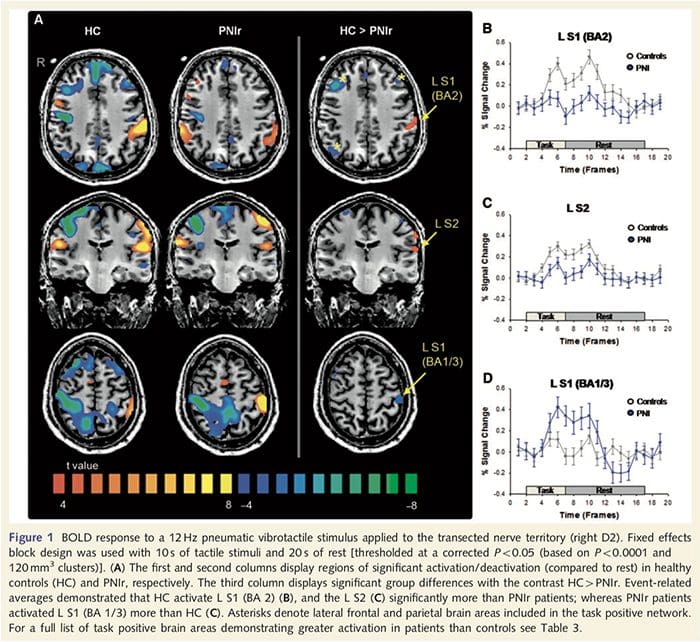
Peta fungsional MRI dihitung dari 11 pasien PNIr dengan transeksi saraf median kanan (pasien dengan transeksi saraf ulnaris dikeluarkan dari analisis ini) dan 11 kontrol sehat yang sesuai usia dan jenis kelamin. Dari Gambar. 1A, jelas bahwa pasien PNIr memiliki aktivasi yang jauh lebih sedikit, dibandingkan dengan kontrol yang sehat, di wilayah S1 yang sesuai dengan area Brodmann 2 (BA2) (Talairach dan Tournoux, 1988) dan S2 (lihat Tabel 3 untuk detailnya) . Respons terkait peristiwa rata-rata dari wilayah yang menarik ini menyoroti respons BOLD yang dilemahkan dalam BA2 kiri pasien dan S2 kiri (Gbr. 1B dan C, masing-masing). Anehnya, stimulasi vibrotaktil pada pasien mengaktifkan bagian yang lebih superior dari girus pasca-sentral (mungkin BA1 / 3) (Talairach dan Tournoux, 1988) (Gbr. 1A dan Tabel 3). Rata-rata terkait peristiwa (Gbr. 1D) menunjukkan bahwa kontrol yang sehat memiliki aktivasi minimal di wilayah ini. Lebih lanjut, pasien memiliki aktivasi yang lebih signifikan di daerah otak yang secara kolektif dikenal sebagai jaringan tugas positif (tanda bintang pada Gambar 1). Lihat Tabel 3 untuk daftar lengkap area otak positif tugas yang diaktifkan. Jaringan ini mencakup korteks prefrontal lateral, parietal lateral, premotor dan temporal inferior (Tabel 3): area otak yang diaktifkan selama kinerja tugas yang menuntut perhatian dan ditekan atau tidak aktif selama istirahat atau tugas yang tidak menantang secara kognitif atau perhatian (Fox et. al., 2005; DeLuca et al., 2006; Seminowicz dan Davis 2007).
Mengurangi Masalah Abu-abu Di Korteks Somatosensori Primer Berkorelasi Dengan Pemulihan Sensorik
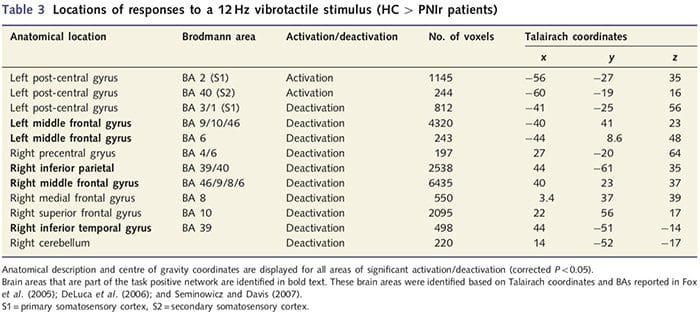
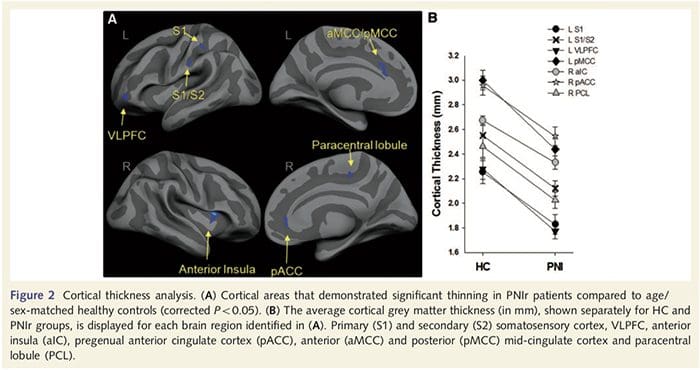
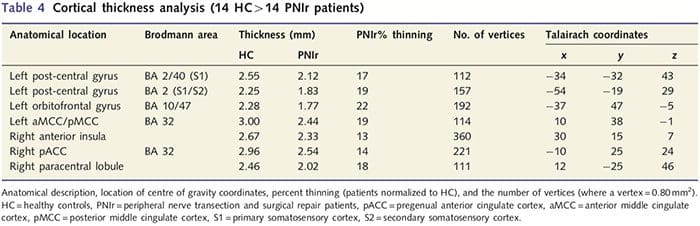
Analisis ketebalan kortikal pada 14 pasien dan 14 kontrol sehat dengan usia/jenis kelamin mengungkapkan beberapa lokus penipisan kortikal yang signifikan pada kelompok PNIr (Gbr. 2 dan Tabel 4). Secara khusus, pasien mengalami penurunan 13%-22% dalam ketebalan kortikal di kiri (kontralesi) S1, S2, girus cingulate anterior pregenual, korteks prefrontal ventrolateral dan insula anterior kanan, gyrus cingulate tengah anterior/posterior dan lobulus paracentral. Menariknya, lokasi penipisan materi abu-abu dalam gyrus pasca-pusat bertepatan dengan daerah BOLD yang berkurang setelah stimulasi vibrotaktil (Tabel 4). Karena kami memiliki pengetahuan sebelumnya tentang defisit sensorik pasien dan waktu pemulihan (yaitu waktu sejak perbaikan bedah mikro), kami selanjutnya bertanya apakah ketebalan kortikal pasien di girus pasca-sentral berkorelasi dengan ambang deteksi mekanis dan getaran sensorik mereka, atau dengan Waktu Pemulihan. Analisis ini mengungkapkan korelasi negatif antara ketebalan kortikal dan ambang deteksi getaran di wilayah yang mencakup BA1/2 dan S2 (P50.001, r=?0.80 dan ?0.91, untuk BA1/2 dan S2, masing-masing; Gambar 3 dan Tabel 5 ). Selain itu, ambang deteksi mekanis juga berkorelasi negatif dengan ketebalan kortikal di wilayah BA2 yang sedikit lebih unggul dan wilayah S2 yang sama (P50.001, r = -0.83 dan -0.85, untuk BA2 dan S2, masing-masing; Gbr. 3 dan Tabel 5). Namun, kami tidak mengidentifikasi hubungan yang signifikan antara waktu pemulihan dan ketebalan kortikal. Oleh karena itu, penipisan kortikal girus pasca-pusat dikaitkan dengan defisit sensorik yang lebih parah. Namun, kami tidak mengidentifikasi hubungan yang signifikan antara waktu pemulihan dan ketebalan kortikal. Sekali lagi, ada korespondensi antara penipisan kortikal di daerah yang berkorelasi negatif dengan rangsangan getaran dan daerah yang menunjukkan kelainan grup fMRI dan CTA.
Kelainan Matter Putih Setelah Transeksi Saraf
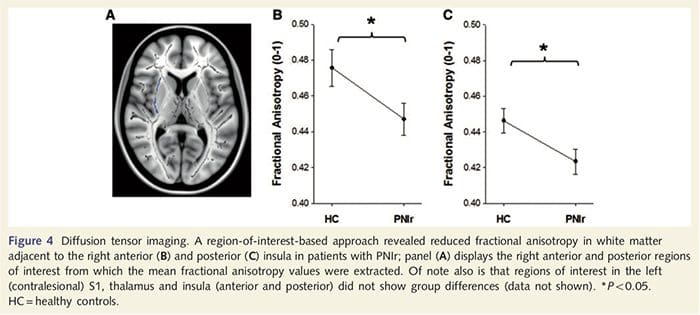
Untuk menilai integritas white matter kami menggunakan pendekatan area of interest untuk menguji perbedaan kelompok materi putih berdasarkan hipotesis-hipotesis apriori. Daerah yang diminati terbatas pada materi putih di sekitar dan memberi makan ke SxNUMX kontralesional dan talamus. Selain itu, kami juga menarik daerah yang tertarik pada materi putih yang berdekatan dengan kiri dan kanan, insula anterior dan posterior. Insula dipilih karena terlibat dalam pengolahan somatosensori dan karena kami mengidentifikasi materi abu-abu berkurang di kanan anterior insular dengan CTA. Pendekatan daerah minat ini mengungkapkan bahwa pasien telah secara signifikan mengurangi nilai-nilai anisotropi pecahan materi putih (MANOVA termasuk semua enam wilayah minat) yang berdekatan dengan anterior kanan [F (1, 1) = 26, P = 4.39; Gambar. 0.046A] dan insula posterior [F (4, 1) = 26, P = 5.55; Gambar. 0.026B], tetapi tidak ada perbedaan kelompok dalam materi putih yang bersebelahan dengan insula kiri (insula anterior sebelah kiri: P = 4; insula posterior kiri: P = 0.51), thalamus (P = 0.26) atau S0.46 (P = 1 ).
 Diskusi
Diskusi
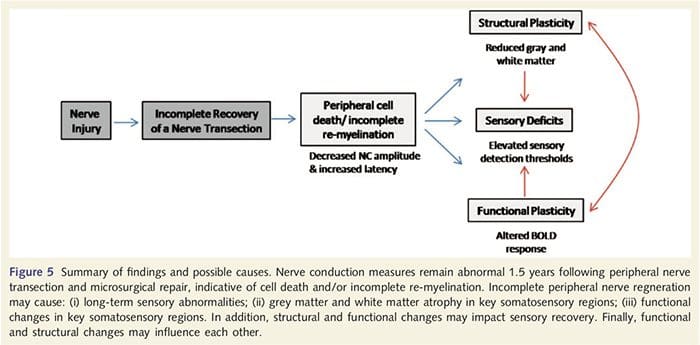
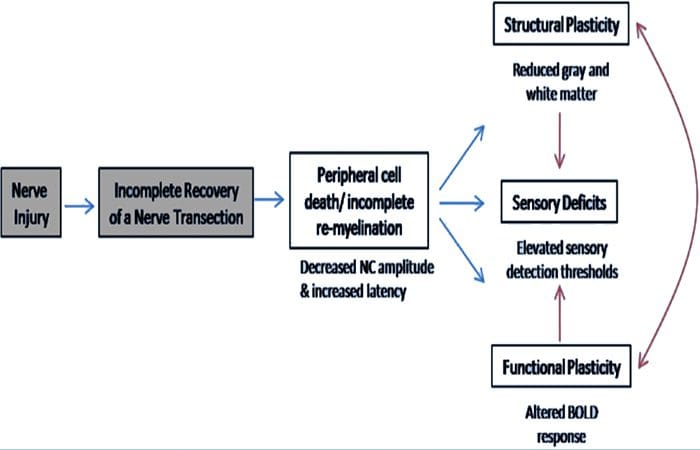
Di sini, kami telah menunjukkan untuk pertama kalinya bahwa ada plastisitas fungsional dan kelainan struktural materi abu-abu dan putih di beberapa daerah kortikal setelah transeksi saraf perifer ekstremitas atas dan perbaikan bedah. Plastisitas ini mungkin timbul dari regenerasi saraf perifer yang tidak lengkap (kematian sel perifer dan / atau mielinasi ulang yang tidak lengkap), karena ukuran konduksi saraf pada pasien ini menunjukkan kelainan berat. Selain itu, data kami menunjukkan bahwa penurunan tanggapan fMRI vibrotaktile-membangkitkan dalam gyrus pasca-sentral sesuai dengan penipisan materi abu-abu dalam kelompok pasien. Hasil ini menunjukkan bahwa pengurangan respon BOLD dapat difasilitasi oleh pengurangan materi abu-abu kortikal dan / atau penurunan input aferen ke gyrus pasca-sentral. Selain itu, ketebalan kortikal dalam bagian-bagian yang sama dari gyrus pasca-sentral berkorelasi negatif dengan ukuran perilaku fungsi somatosensori. Artinya, peningkatan defisit somatosensor berkorelasi dengan korteks yang lebih tipis; keduanya mungkin terkait dengan input aferen. Secara bersama-sama, data kami menunjukkan bahwa regenerasi saraf perifer yang tidak lengkap berkontribusi terhadap gangguan somatosensori, atrofi materi abu-abu kortikal dan mengurangi aktivasi fMRI (lihat Gambar. 5 untuk ringkasan temuan ini).
Telah diketahui bahwa plastisitas kortikal setelah transeksi saraf perifer dan perbaikan bedah dapat terjadi di seluruh CNS pada primata non-manusia (Kaas, 1991). Plastisitas ini dianggap karena pembukaan dari sinapsis diam sebelumnya atau tunas aksonal ke wilayah yang tuli (Wall et al., 1986; Florence dan Kaas, 1995). Dalam model primata, 1 tahun setelah transeksi saraf dan perbaikan bedah, korteks denervasi ditandai oleh representasi yang tidak lengkap dan tidak teratur dari saraf yang diregenerasi dan berdekatan (utuh). Representasi tambal sulam ini dikaitkan dengan regenerasi perifer yang tidak lengkap yang menghasilkan pemulihan parsial dari ruang kortikal denervasi (Kaas, 1991). Untuk menilai sejauh mana regenerasi perifer dalam populasi pasien kami, kami melakukan studi konduksi saraf sensorik dan motorik di seluruh area yang ditransleksi. Hasil konduksi saraf kami menunjukkan bahwa pasien PNIr mengalami penurunan amplitudo secara signifikan dan peningkatan latensi pada saraf sensorik dan motorik dibandingkan dengan sisi mereka yang tidak tertransmisikan. Amplitudo yang menurun dikombinasikan dengan peningkatan latensi merupakan indikasi hilangnya serat perifer (mis. Kematian sel) dan / atau kelainan reeleksiasi abnormal atau tidak lengkap setelah transeksi (Kimura, 1984). Selain itu, sudah diketahui bahwa antara 20% dan 50% dari neuron ganglion akar dorsal mati setelah transeksi saraf (Liss et al., 1996). Dengan demikian, kematian sel aferen dan regenerasi tidak lengkap dapat menyebabkan penurunan masukan aferen ke korteks, yang dapat menyebabkan defisit sensorik yang sedang berlangsung dan penurunan respons BOLD di BA2 dan S2. Lebih jauh lagi, masukan aferen yang berkurang ini juga dapat menjelaskan penipisan kortikal yang kami amati di daerah korteks yang sama. Pencabutan sensorik telah terbukti menyebabkan degenerasi trans-neuronal di beberapa daerah CNS, termasuk tanduk dorsal berikut bagian saraf sciatic (Knyihar-Csillik et al., 1989), dan mungkin melibatkan neuron orde kedua dan ketiga (Powell dan Erulkar, 1962). Degenerasi transneuronal dicirikan oleh penyusutan sel dan dianggap berhubungan dengan penurunan, atau tidak ada, input aferen (Knyihar-Csillik et al., 1989). Dengan demikian, kehilangan materi abu-abu kortikal (atau atrofi) juga bisa langsung terkait dengan penurunan masukan aferen.
 Kami juga menunjukkan peningkatan aktivasi di gyrus pasca-pusat di wilayah yang sesuai dengan BA1 / 3 (Talairach dan Tournoux, 1988). Elektrofisiologi, penelusuran anatomi dan penelitian neuroimaging telah menetapkan bahwa untuk sebagian besar aferen mekanoreceptive kulit tujuan kortikal pertama adalah BA1 dan BA3b. Area otak cytoarchitectonic ini masing-masing memiliki peta tubuh somatotopic dengan bidang reseptif kecil. Selain itu, area ini merespon banyak fitur informasi taktil, seperti tekstur dan kekasaran, kecepatan dan kelengkungan rangsangan (Bodegard et al., 2001). Studi fMRI telah menunjukkan bahwa aktivitas dalam korteks somatosensori dipengaruhi oleh perhatian sedemikian rupa sehingga tanggapan fMRI terhadap rangsangan taktil di S1 meningkat ketika subjek menghadiri stimulus taktil, tetapi dilemahkan ketika subjek terganggu (Arthurs et al., 2004; Porro et al., 2004). Selanjutnya, pasien kami mengaktifkan jaringan area otak yang dikenal sebagai jaringan tugas positif (DeLuca et al., 2006) lebih dari kontrol yang sehat. Area otak ini diaktifkan selama proses yang menuntut perhatian (Fox et al., 2005; Seminowicz dan Davis, 2007). Bersama-sama, temuan ini menyiratkan bahwa pasien harus memperhatikan stimulus lebih dari kontrol karena gangguan masukan sensorik mereka. Peningkatan perhatian ini juga dapat menjelaskan peningkatan aktivasi di BA1 / 3b. Tentu saja, peningkatan aktivasi di BA1 / 3b juga dapat mencerminkan plastisitas yang tidak terkait dengan beban attentional.
Kami juga menunjukkan peningkatan aktivasi di gyrus pasca-pusat di wilayah yang sesuai dengan BA1 / 3 (Talairach dan Tournoux, 1988). Elektrofisiologi, penelusuran anatomi dan penelitian neuroimaging telah menetapkan bahwa untuk sebagian besar aferen mekanoreceptive kulit tujuan kortikal pertama adalah BA1 dan BA3b. Area otak cytoarchitectonic ini masing-masing memiliki peta tubuh somatotopic dengan bidang reseptif kecil. Selain itu, area ini merespon banyak fitur informasi taktil, seperti tekstur dan kekasaran, kecepatan dan kelengkungan rangsangan (Bodegard et al., 2001). Studi fMRI telah menunjukkan bahwa aktivitas dalam korteks somatosensori dipengaruhi oleh perhatian sedemikian rupa sehingga tanggapan fMRI terhadap rangsangan taktil di S1 meningkat ketika subjek menghadiri stimulus taktil, tetapi dilemahkan ketika subjek terganggu (Arthurs et al., 2004; Porro et al., 2004). Selanjutnya, pasien kami mengaktifkan jaringan area otak yang dikenal sebagai jaringan tugas positif (DeLuca et al., 2006) lebih dari kontrol yang sehat. Area otak ini diaktifkan selama proses yang menuntut perhatian (Fox et al., 2005; Seminowicz dan Davis, 2007). Bersama-sama, temuan ini menyiratkan bahwa pasien harus memperhatikan stimulus lebih dari kontrol karena gangguan masukan sensorik mereka. Peningkatan perhatian ini juga dapat menjelaskan peningkatan aktivasi di BA1 / 3b. Tentu saja, peningkatan aktivasi di BA1 / 3b juga dapat mencerminkan plastisitas yang tidak terkait dengan beban attentional.
 BA2 dan S2 keduanya menerima proyeksi dari BA1 / 3b dan juga dari bagian yang berbeda dari kompleks thalamus ventroposterior (Pons et al., 1985; Friedman dan Murray, 1986). Keduanya otak daerah memiliki bidang reseptif yang besar, seringkali multi-digit (BA2) atau bilateral (S2) (Pons et al., 1985; Iwamura et al., 2002). Berdasarkan proyeksi anatomis dan sifat respons neuron, pemrosesan hierarki informasi taktil telah dibuktikan dari BA1 / 3b hingga BA 2 (Kaas et al., 2002). Selain itu, studi elektrofisiologi pada kera (Pons et al., 1987) dan data magnetoence-phalography yang diperoleh pada manusia, menunjukkan bahwa pemrosesan serial input taktil terjadi dari S1 ke S2 pada primata yang lebih tinggi (Frot dan Mauguiere 1999; Disbrow et al. ., 2001). Beberapa penelitian telah menunjukkan bahwa BA2 secara istimewa diaktifkan oleh bentuk dan kelengkungan (Bodegard et al., 2001), sedangkan S2 mungkin terlibat dalam pembelajaran taktil (Ridley dan Ettlinger 1976; Murray dan Mishkin, 1984), mendukung gagasan bahwa area otak ini terlibat dalam pemrosesan somatosensori tingkat tinggi. Penilaian psikofisik kami menunjukkan bahwa pasien secara signifikan terganggu saat mendeteksi rangsangan sentuhan sederhana, dan dalam tes Identifikasi Tekstur Bentuk 1.5 tahun setelah operasi (Taylor et al., 2008a). Tes terakhir ini menilai kemampuan pasien untuk mengenali karakteristik suatu objek sambil secara aktif mengeksplorasi bentuk atau tekstur, membutuhkan integrasi informasi sensorik di seluruh wilayah tubuh (Rosen dan Lundborg, 1998). Secara keseluruhan, satu interpretasi dari data kami adalah bahwa pasien PNIr lebih memperhatikan stimulus vibrotaktil, yang mengarah ke peningkatan aktivasi jaringan positif tugas dan BA1 / 3. Namun, pada pasien ini, data kami menyiratkan bahwa area pemrosesan tingkat tinggi, seperti BA2 dan S2, tidak menerima informasi taktil, yang, pada gilirannya, dapat mengakibatkan penipisan kortikal dan pengurangan respons BOLD.
BA2 dan S2 keduanya menerima proyeksi dari BA1 / 3b dan juga dari bagian yang berbeda dari kompleks thalamus ventroposterior (Pons et al., 1985; Friedman dan Murray, 1986). Keduanya otak daerah memiliki bidang reseptif yang besar, seringkali multi-digit (BA2) atau bilateral (S2) (Pons et al., 1985; Iwamura et al., 2002). Berdasarkan proyeksi anatomis dan sifat respons neuron, pemrosesan hierarki informasi taktil telah dibuktikan dari BA1 / 3b hingga BA 2 (Kaas et al., 2002). Selain itu, studi elektrofisiologi pada kera (Pons et al., 1987) dan data magnetoence-phalography yang diperoleh pada manusia, menunjukkan bahwa pemrosesan serial input taktil terjadi dari S1 ke S2 pada primata yang lebih tinggi (Frot dan Mauguiere 1999; Disbrow et al. ., 2001). Beberapa penelitian telah menunjukkan bahwa BA2 secara istimewa diaktifkan oleh bentuk dan kelengkungan (Bodegard et al., 2001), sedangkan S2 mungkin terlibat dalam pembelajaran taktil (Ridley dan Ettlinger 1976; Murray dan Mishkin, 1984), mendukung gagasan bahwa area otak ini terlibat dalam pemrosesan somatosensori tingkat tinggi. Penilaian psikofisik kami menunjukkan bahwa pasien secara signifikan terganggu saat mendeteksi rangsangan sentuhan sederhana, dan dalam tes Identifikasi Tekstur Bentuk 1.5 tahun setelah operasi (Taylor et al., 2008a). Tes terakhir ini menilai kemampuan pasien untuk mengenali karakteristik suatu objek sambil secara aktif mengeksplorasi bentuk atau tekstur, membutuhkan integrasi informasi sensorik di seluruh wilayah tubuh (Rosen dan Lundborg, 1998). Secara keseluruhan, satu interpretasi dari data kami adalah bahwa pasien PNIr lebih memperhatikan stimulus vibrotaktil, yang mengarah ke peningkatan aktivasi jaringan positif tugas dan BA1 / 3. Namun, pada pasien ini, data kami menyiratkan bahwa area pemrosesan tingkat tinggi, seperti BA2 dan S2, tidak menerima informasi taktil, yang, pada gilirannya, dapat mengakibatkan penipisan kortikal dan pengurangan respons BOLD.
Insula ini diduga berperan dalam mengintegrasikan informasi multimodal yang penting untuk fungsi sensorimotor, emosional, allostatik / homeostatik dan kognitif (Devinsky dkk., 1995; Critchley, 2004; Craig, 2008) dan telah ditunjuk sebagai korteks sensorik limbik (Craig , 2008). Beberapa penelitian telah melaporkan aktivasi insular dalam menanggapi rangsangan taktil (Gelnar et al., 1998;
Downar et al., 2002) dan studi penelusuran anatomi pada primata telah menunjukkan bahwa insula terhubung secara resipien ke lobus frontal, parietal dan temporal (Agustinus, 1996). Pada pasien kami, insula anterior kanan adalah satu-satunya daerah kortikal yang menunjukkan penipisan kortikal yang signifikan dalam hubungannya dengan penurunan nilai anisotropi fraksional pada materi putih yang berdekatan, menunjukkan bahwa penipisan kortikal dalam daerah ini dikaitkan dengan hilangnya serat yang memproyeksikan ke atau dari struktur ini. Insula anterior kanan telah terlibat dalam interoception karena terletak untuk mengintegrasikan input homeostatik dari tubuh dengan kondisi motivasi, emosional dan sosial (Craig, 2008). Selanjutnya, Critchley dkk. (2004) melaporkan korelasi antara kemampuan interoceptive dan volume abu-abu dari insula anterior kanan. Mengingat temuan kami bahwa pasien mengalami penurunan materi abu-abu di insula anterior kanan, akan menarik untuk menilai kemampuan interoceptive setelah cedera saraf perifer dalam studi masa depan.
Secara bersama-sama, kami telah menunjukkan untuk pertama kalinya bahwa perubahan fungsional dan struktural hadir dalam manusia korteks 1.5 tahun setelah transeksi lengkap dari saraf perifer ekstremitas atas yang diperbaiki secara microsurgical. Selain itu, pengukuran konduksi saraf menunjukkan regenerasi perifer yang tidak sempurna pada pasien ini. Selanjutnya, kami menunjukkan bahwa ketebalan kortikal terkait dengan pengukuran psikofisik pemulihan, dalam korteks yang lebih tipis dalam BA2 dan S2 dikaitkan dengan fungsi somatosensori yang lebih buruk. Data ini menunjukkan bahwa pembentukan kembali peta aktivasi fungsional normal secara langsung terkait dengan regenerasi sukses aferen periferal.
Keri S. Taylor, 1,2 Dimitri J. Anastakis2,3,4 dan Karen D. Davis1,2,3
1 Divisi Otak, Pencitraan dan Perilaku Ilmu Saraf Sistem, Toronto Western Research Institute, University Health Network, Toronto, Kanada M5T258
2 Institute of Medical Science, Universitas Toronto, Kanada
3 Department of Surgery, University of Toronto, Kanada
Pusat Sumber Studi Klinis 4, Institut Penelitian Barat Toronto, Jaringan Kesehatan Universitas, Toronto, Kanada M5T2S8
Korespondensi ke: Karen D. Davis, Ph.D.,
Divisi Otak, Pencitraan dan Perilaku Ilmu Saraf Sistem, Toronto Western Research Institute,
Rumah Sakit Toronto Barat,
Jaringan Kesehatan Universitas,
Ruang MP14-306, 399 Bathurst Street,
Toronto, Ontario,
Kanada M5T 2S8
E-mail: kdavis@uhnres.utoronto.ca
Ucapan Terima Kasih
Para penulis mengucapkan terima kasih kepada Bapak Geoff Pope, Dr. Adrian Crawley, Tn. Eugene Hlasny, dan Mr. Keith Ta untuk bantuan teknis ahli. Para penulis ingin mengucapkan terima kasih kepada Dr. Peter Ashby dan Mr. Freddy Paiz dari Klinik EMG Toronto Rumah Sakit Barat untuk melakukan tes konduksi saraf dan untuk menyediakan penilaian ahli atas temuan. Para penulis juga berterima kasih kepada Drs Dvali, Binhammer, Fialkov dan Antonyshyn untuk berkolaborasi dengan proyek ini. Dr. Davis adalah Ketua Penelitian Kanada dalam Otak dan Perilaku (CIHR MOP 53304).
Pendanaan
The Physicians Services Incorporated dan hibah benih bersama dari Pusat Studi Nyeri Universitas Toronto / AstraZeneca.
Materi tambahan
Materi tambahan tersedia di Brain online.
kosong
Referensi:
Apkarian AV, Sosa Y, Sonty S, dkk. Nyeri punggung kronis dikaitkan dengan
penurunan kepadatan materi abu-abu prefrontal dan talamus. J Neurosci
2004; 24: 10410�5.
Arthurs OJ, Johansen-Berg H, Matthews PM, Boniface SJ. Perhatian
secara berbeda memodulasi kopling fMRI BOLD dan membangkitkan
amplitudo sinyal potensial di korteks somatosensori manusia7.
Exp Brain Res 2004; 157: 269�74.
Agustinus JR. Sirkuit dan aspek fungsional lobus insular
pada primata termasuk manusia. Brain Res Brain Res Rev 1996; 22:
229�44.
Beaulieu C. Dasar difusi air anisotropik di saraf
sistem – tinjauan teknis. NMR Biomed 2002; 15: 435�55.
Bodegard A, Geyer S, Grefkes C, Zilles K, Roland PE. hierarkis
pemrosesan bentuk taktil di otak manusia. Neuron 2001; 31:
317�28.
Craig AD. Interoception dan Emosi: Sebuah Perspektif Neuroanatomis.
Dalam: Lewis M, Haviland-Jones J, Barrett L, editor. Buku pegangan dari
emosi. New York: Pers Guildford; 2008. hal. 272�87.
Critchley HD. Korteks manusia merespons tantangan interoseptif.
Proc Natl Acad Sci USA 2004; 101: 6333�4.
Critchley HD, Wiens S, Rotshtein P, Ohman A, Dolan RJ. Sistem saraf
mendukung kesadaran interoseptif. Nat Neurosci 2004; 7: 189�95.
Dale AM, Fischl B, Sereno MI. Analisis berbasis permukaan kortikal. I. Segmentasi
dan rekonstruksi permukaan. Neurocitra 1999; 9: 179�94.
Davis KD, Pope G, Chen J, Kwan CL, Crawley AP, Diamant NE. kortikal
penipisan di IBS: implikasi untuk homeostatis, perhatian, dan rasa sakit
pengolahan. Neurologi 2008; 70: 153�4.
DeLuca M, Beckmann CF, De SN, Matthews PM, Smith SM. fMRI istirahat
jaringan negara mendefinisikan mode interaksi jarak jauh yang berbeda
di otak manusia. Neuroimage 2006; 29: 1359�67.
Devinsky O, Morrell MJ, Vogt BA. Kontribusi cingulate anterior
korteks untuk perilaku. Otak 1995; 118 (Pt 1): 279�306.
Disbrow E, Roberts T, Poeppel D, Krubitzer L. Bukti untuk interhemispheric
pemrosesan input dari tangan di S2 manusia
dan PV. J Neurofisiol 2001; 85: 2236�44.
Downar J, Crawley AP, Mikulis DJ, Davis KD. Jaringan kortikal sensitif
untuk merangsang arti-penting dalam konteks perilaku netral di berbagai
modalitas sensorik. J Neurofisiologi 2002; 87: 615�20.
Draganski B, Moser T, Lummel N, dkk. Penurunan abu-abu thalamic
masalah setelah amputasi anggota badan. Neuroimage 2006; 31: 951�7.
Fischl B, Dale AM. Mengukur ketebalan korteks serebral manusia
dari gambar resonansi magnetik. Proc Natl Acad Sci USA 2000; 97:
11050�5.
Fischl B, Sereno MI, Dale AM. Analisis berbasis permukaan kortikal. II:
Inflasi, perataan, dan sistem koordinat berbasis permukaan.
Neurocitra 1999a; 9: 195�207.
Fischl B, Sereno MI, Tootell RB, Dale AM. Intersubjek resolusi tinggi
rata-rata dan sistem koordinat untuk permukaan kortikal. Otak Hum
Peta 1999b; 8: 272�84.
Florence SL, Kaas JH. Reorganisasi skala besar di berbagai tingkat
Jalur somatosensori mengikuti terapi amputasi tangan
di monyet. J Neurosci 1995; 15: 8083�95.
Fox MD, Snyder AZ, Vincent JL, Corbetta M, Van E, Raichle ME. Itu
otak manusia secara intrinsik diatur menjadi dinamis, antikorelasi
jaringan fungsional. Proc Natl Acad Sci USA 2005; 102: 9673�9678.
Friedman DP, Murray EA. Konektivitas thalamic yang kedua
daerah somatosensori dan bidang somatosensori tetangganya
sulkus lateral kera. J Comp Neurol 1986; 252: 348�73.
Frot M, Mauguiere F. Waktu dan distribusi spasial somatosensori
tanggapan dicatat di tepi atas fisura sylvian (area SII) di
manusia. Korteks Serebrum 1999; 9: 854�63.
Geha PY, Baliki MN, Harden RN, Bauer WR, Parrish TB, Apkarian AV.
Otak pada nyeri CRPS kronis: materi abu-abu-putih abnormal
interaksi di daerah emosional dan otonom. Neuron 2008; 60:
570�81.
Gelnar PA, Krauss BR, Szeverenyi NM, Apkarian AV. Representasi ujung jari
di korteks somatosensori manusia: sebuah studi fMRI.
Neurocitra 1998; 7: 261�83.
Hickmott PW, Steen PA. Perubahan skala besar dalam struktur dendritik
selama reorganisasi korteks somatosensori dewasa. Nat Neurosci
2005; 8: 140�42.
Iwamura Y, Tanaka M, Iriki A, Taoka M, Toda T. Pemrosesan
sinyal taktil dan kinestetik dari sisi bilateral tubuh di
gyrus postcentral monyet terjaga. Perilaku Otak Res 2002; 135:
185�90.
Jaquet JB, Luijsterburg AJ, Kalmijn S, Kuypers PD, Hofman A, Hovius SE.
Cedera saraf median, ulnaris, dan gabungan median-ulnaris: fungsional
hasil dan kembali ke produktivitas. J Trauma 2001; 51: 687�92.
Jurkiewicz MT, Crawley AP, Verrier MC, Fehlings MG, Mikulis DJ.
Atrofi kortikal somatosensori setelah cedera tulang belakang: berbasis voxel
studi morfometri. Neurologi 2006; 66: 762�4.
Kaas JH. Plastisitas peta sensorik dan motorik pada mamalia dewasa. annu
Pdt Neurosci 1991; 14: 137�67.
Kaas JH, Jain N, Qi HX. Organisasi sistem somatosensori dalam
primata. Dalam: Nelson RJ, editor. Sistem somatosensori.
Washington, DC: CRC Press; 2002. hal. 1�25.
Kimura J. Elektrodiagnosis pada penyakit saraf dan otot: prinsip
dan praktek. Oxford: Pers Universitas Oxford; 2001.
Kimura J. Prinsip dan perangkap studi konduksi saraf. Ann Neurol
1984; 16: 415�29.
Knyihar-Csillik E, Rakic P, Csillik B. Degenerasi transneuronal di
substansi Rolando dari sumsum tulang belakang primata yang ditimbulkan oleh
atrofi degeneratif transganglionik yang diinduksi axotomy dari pusat
terminal sensorik primer. Res Jaringan Sel 1989; 258: 515�25.
Liss AG, af Ekenstam FW, Wiberg M. Hilangnya neuron di akar dorsal
ganglia setelah transeksi saraf sensorik perifer. Sebuah anatomi
belajar pada monyet. Scand J Plast Reconstr Surg Hand Surg 1996; 30: 1�6.
Gerakan Lotze M, Flor H, Grodd W, Larbig W, Birbaumer N. Phantom
dan rasa sakit. Sebuah studi fMRI pada diamputasi ekstremitas atas. Otak 2001;
124: 2268�77.
Manduch M, Bezuhly M, Anastakis DJ, Crawley AP, Mikulis DJ. Serial
fMRI dari perubahan adaptif di korteks sensorimotor primer berikut
rekonstruksi ibu jari. Neurologi 2002; 59: 1278�81.
Mei A. Nyeri kronis dapat mengubah struktur otak. Sakit 2008;
137: 7�15.
McAllister RM, Gilbert SE, Calder JS, Smith PJ. Epidemiologi dan
manajemen cedera saraf perifer ekstremitas atas di modern
praktek. J Hand Surg (Br) 1996; 21: 4�13.
Murray EA, Mishkin M. Kontribusi relatif dari SII dan area5 terhadap taktil
diskriminasi pada monyet 2. Behav Brain Res 1984; 11: 67�83.
Napadow V, Kettner N, Ryan A, Kwong KK, Audette J, Hui KK.
Plastisitas kortikal somatosensori pada carpal tunnel syndrome-a crosssectional
evaluasi fMRI. Neuroimage 2006; 31: 520�30.
Oldfield RC. Penilaian dan analisis wenangan: Edinburgh
inventaris. Neuropsikologi 1971; 9: 97�113.
Pons TP, Garraghty PE, Cusick CG, Kaas JH. Organisasi somatotopik
area 2 pada kera kera 6. J Comp Neurol 1985; 241: 445�66.
Pons TP, Garraghty PE, Friedman DP, Mishkin M. Bukti fisiologis
untuk pemrosesan serial di korteks somatosensori. Sains 1987; 237:
417�20.
Porro CA, Lui F, Facchin P, Maieron M, Baraldi P. Aktivitas terkait persepsi
dalam sistem somatosensori manusia: resonansi magnetik fungsional
studi pencitraan. Pencitraan Magn Reson 2004; 22: 1539�48.
Powell TP, Erulkar S. Degenerasi sel transneuronal di pendengaran
inti estafet kucing. J Anat 1962; 96: 249�68.
Ridley RM, Ettlinger G. Gangguan pembelajaran taktil dan retensi setelah
penghapusan korteks proyeksi sensorik somatik kedua (SII) di
monyet. Otak Res 1976; 109: 656�60.
Rosen B, Lundborg G. Instrumen gnosis taktil baru dalam sensibilitas
pengujian. J Tangan Ada 1998; 11: 251�7.
Seminowicz DA, Davis KD. Rasa sakit meningkatkan konektivitas fungsional
jaringan otak yang ditimbulkan oleh kinerja tugas kognitif.
J Neurofisiol 2007; 97: 3651�9.
Smith SM, Jenkinson M, Johansen-Berg H, dkk. Spasial berbasis traktat
statistik: analisis voxelwise data difusi multi-subjek.
Neurocitra 2006; 31: 1487�1505.
Talairach J, Tournoux P. Atlas stereotaxic co-planar otak manusia.
New York: Thieme Medical Publishers Inc.; 1988.
Taylor KS, Anastakis DJ, Davis KD. Nyeri kronis setelah perifer
cedera saraf dikaitkan dengan bencana nyeri dan neurotisisme.
Int Ass Stud Pain 2008a; 267.
Taylor KS, Davis KD. Stabilitas fMRI yang berhubungan dengan taktil dan nyeri
aktivasi otak: pemeriksaan ambang batas dan
metode ambang-independen. Hum Brain Peta 2009; 30:
1947�62.
Taylor KS, Seminowicz DA, Davis KD. Dua sistem keadaan istirahat
konektivitas antara insula dan korteks cingulate. Otak Hum
Peta 2008b;DOI:10.1002/hbm.20705.
Wall JT, Kaas JH, Sur M, Nelson RJ, Felleman DJ, Merzenich MM.
Reorganisasi fungsional di area kortikal somatosensori 3b dan 1
monyet dewasa setelah perbaikan saraf median: kemungkinan hubungan dengan
pemulihan sensorik pada manusia. J Neurosci 1986; 6: 218 33.

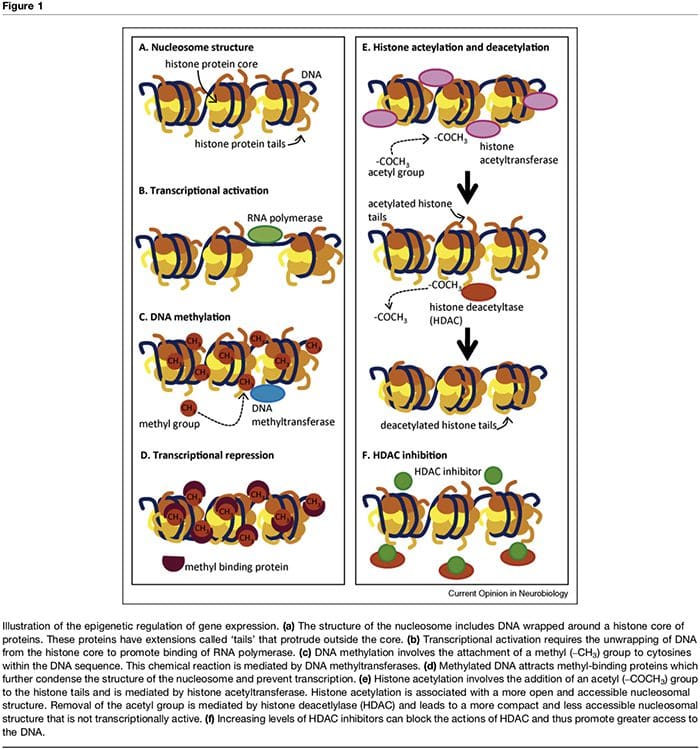




 Saraf kranial I-XII WNL
Saraf kranial I-XII WNL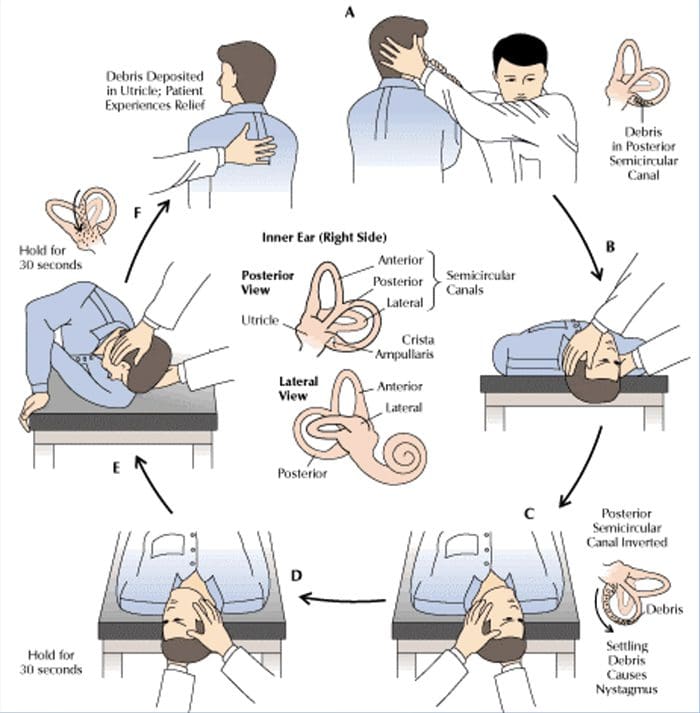 Manuver Reposisi Telapak Posterior Kanan.
Manuver Reposisi Telapak Posterior Kanan. Peningkatan yang ditandai dalam keseimbangan.
Peningkatan yang ditandai dalam keseimbangan.


 APA YANG KAMU LIHAT?
APA YANG KAMU LIHAT? APA YANG KAMU LIHAT?
APA YANG KAMU LIHAT?

 AFFERENTASI DENGAN PERTIMBANGAN METABOLIK
AFFERENTASI DENGAN PERTIMBANGAN METABOLIK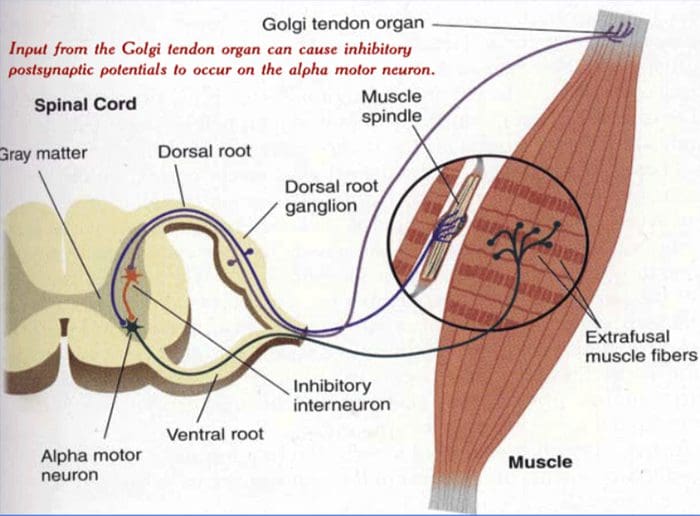
 Di 2011, Aaron kehilangan kedua matanya dalam ledakan IED.
Di 2011, Aaron kehilangan kedua matanya dalam ledakan IED.
 Setelah sembuh dari meningitis:
Setelah sembuh dari meningitis:

 TANTANGAN DITERIMA
TANTANGAN DITERIMA
 Pembaruan dari McKayla:
Pembaruan dari McKayla:


 BEBERAPA TERAPI CERDAS SEDERHANA
BEBERAPA TERAPI CERDAS SEDERHANA





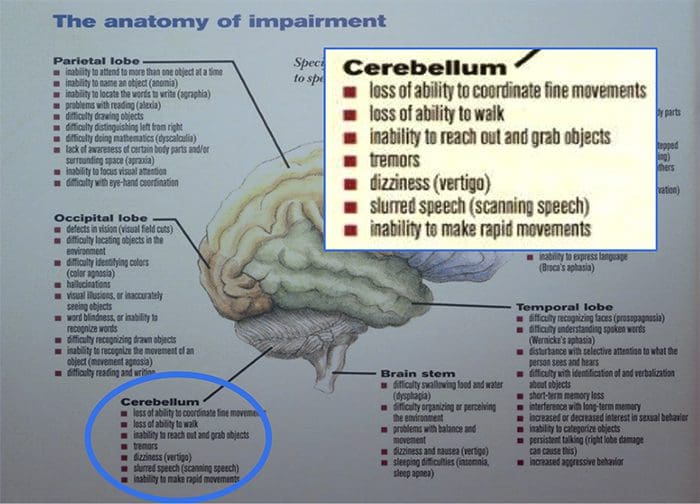
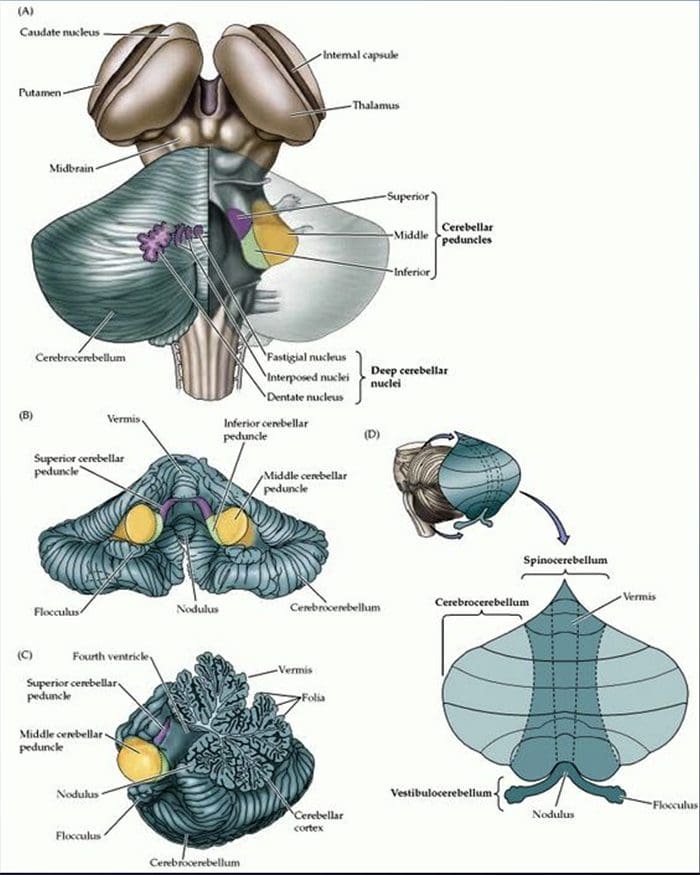
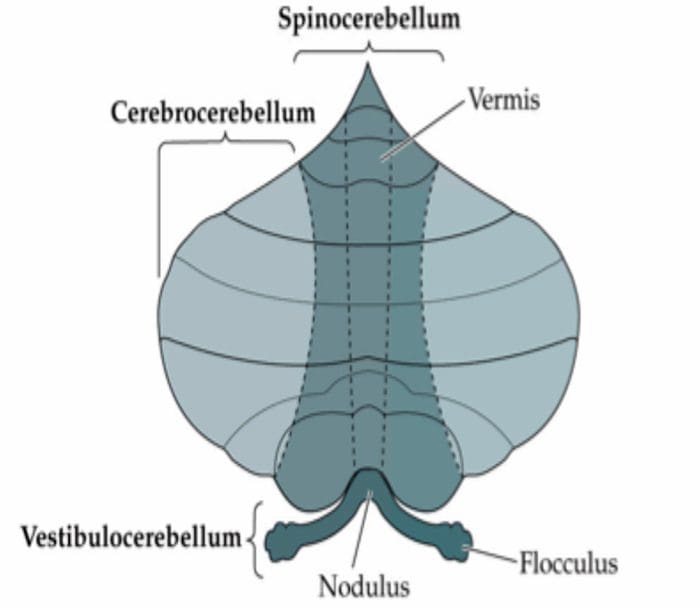 Tanggung Jawab:
Tanggung Jawab:


 APA YANG BISA ANDA LAKUKAN?
APA YANG BISA ANDA LAKUKAN? Mintalah pasien melakukan latihan keseimbangan:
Mintalah pasien melakukan latihan keseimbangan:
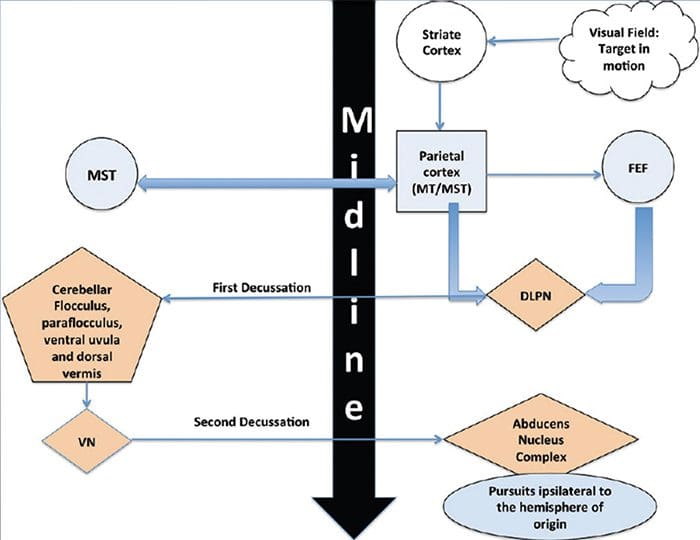
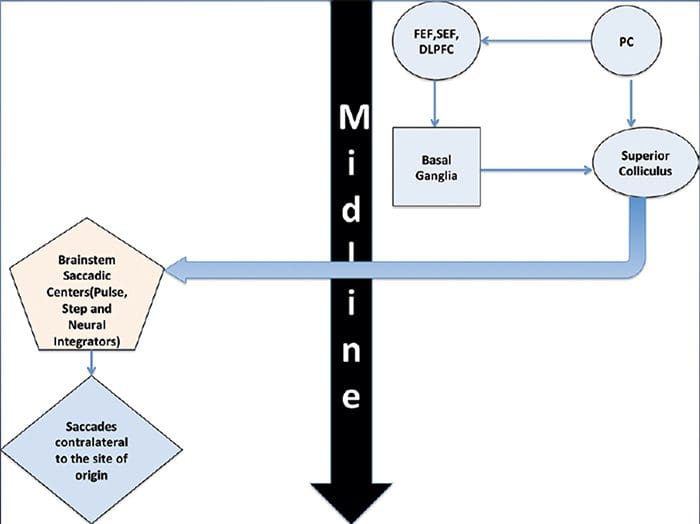
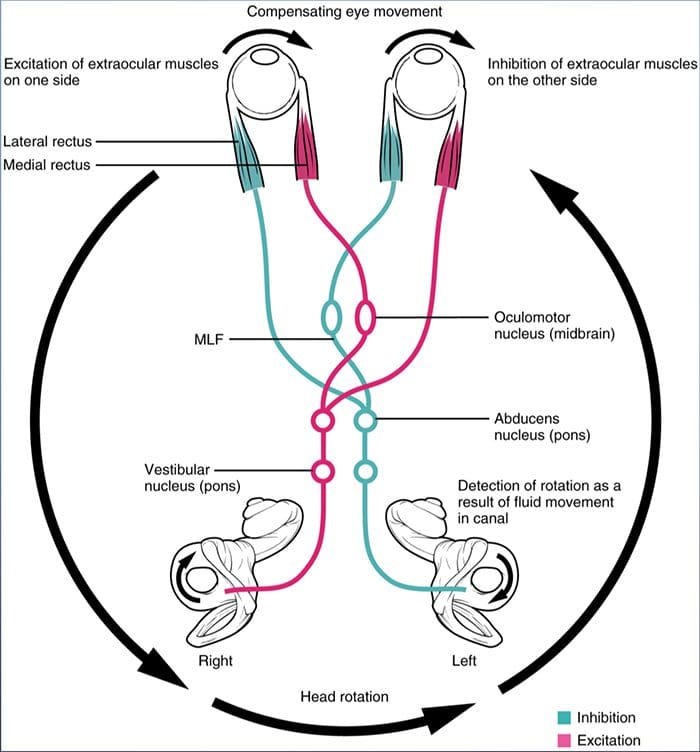
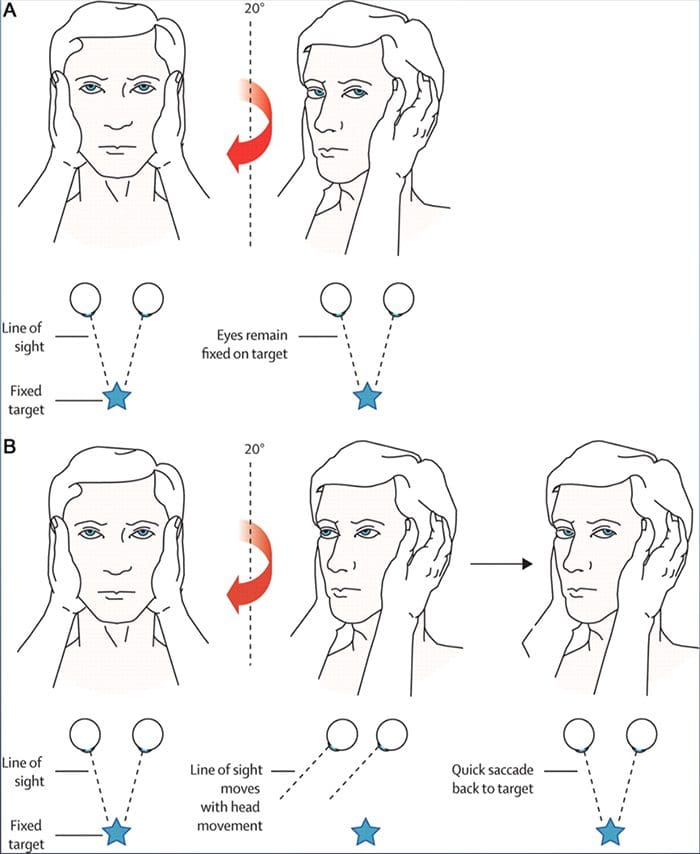
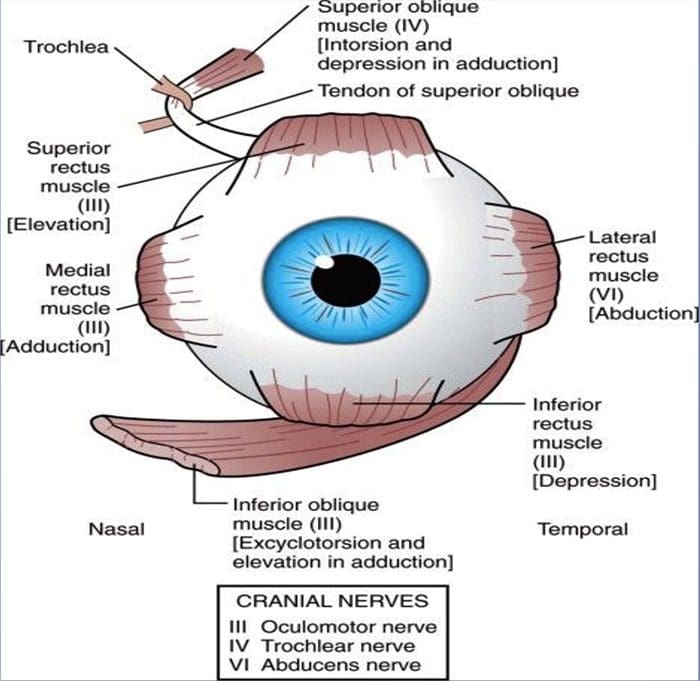
 REVIEW GERAKAN EYE
REVIEW GERAKAN EYE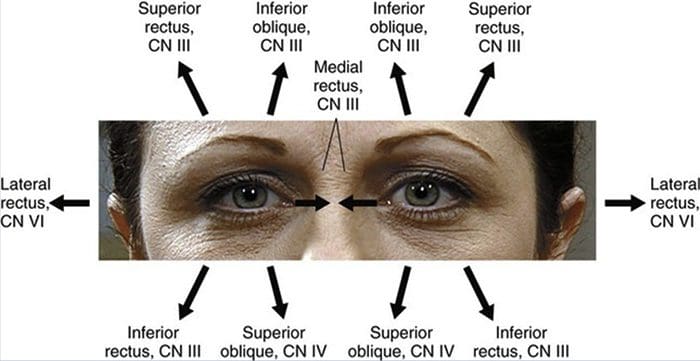
 APA YANG BISA ANDA LAKUKAN?
APA YANG BISA ANDA LAKUKAN? Mintalah pasien melakukan latihan stabilitas pandangan:
Mintalah pasien melakukan latihan stabilitas pandangan:


 APA YANG BISA ANDA LAKUKAN?
APA YANG BISA ANDA LAKUKAN?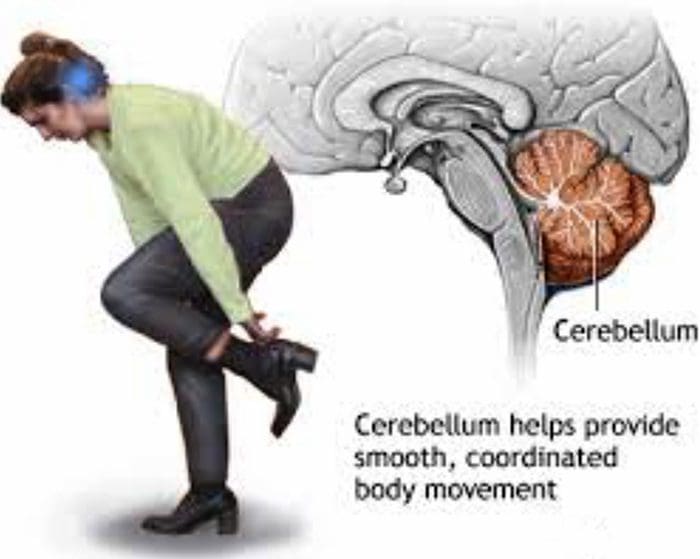 Minta pasien melakukan gerakan terkoordinasi!
Minta pasien melakukan gerakan terkoordinasi!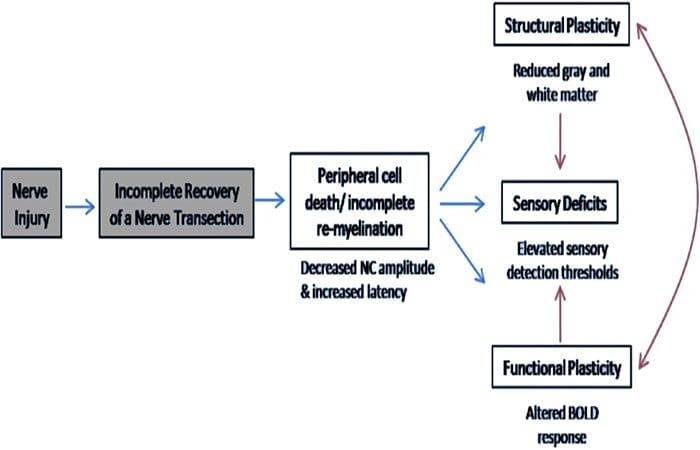

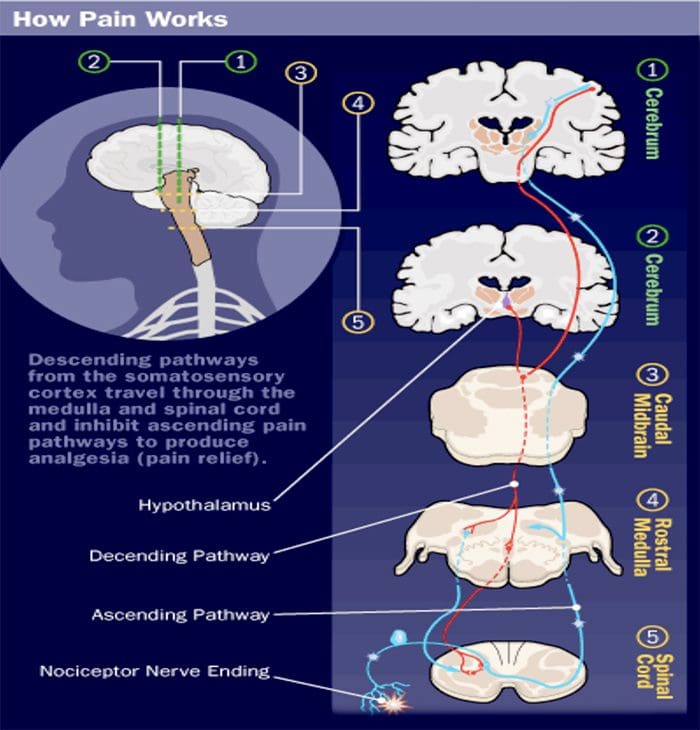
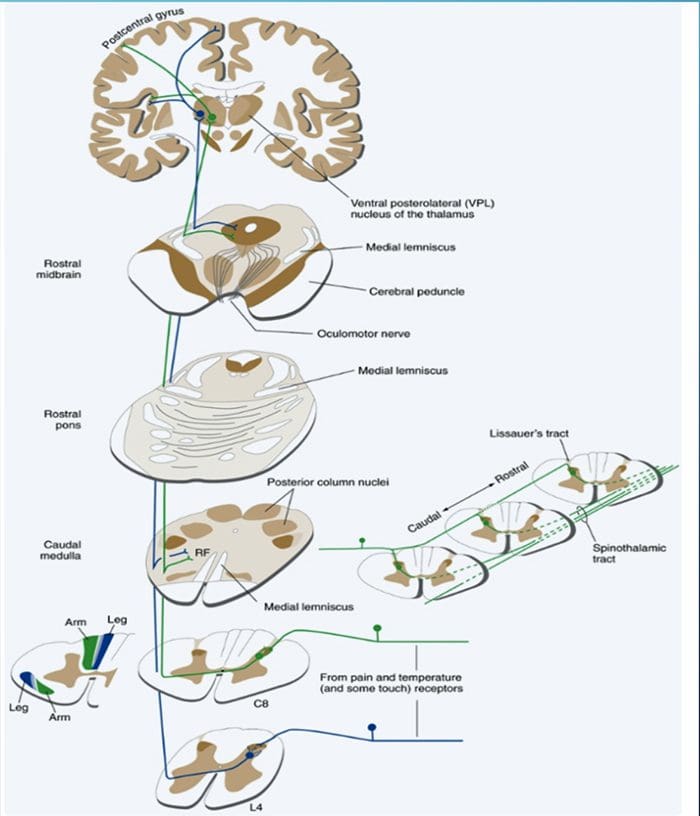
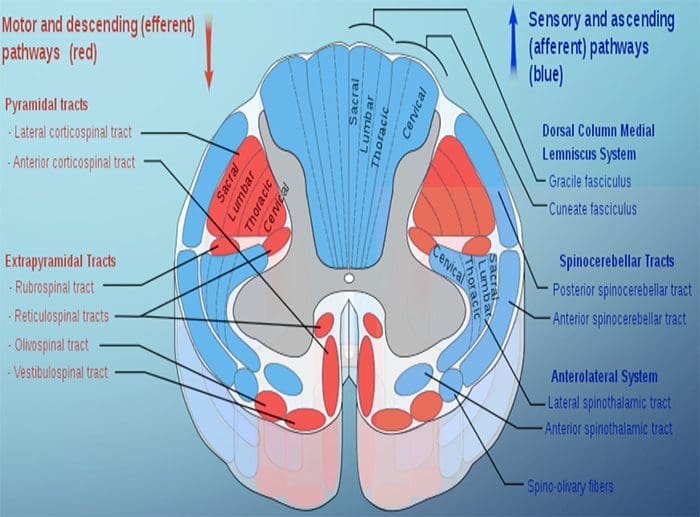
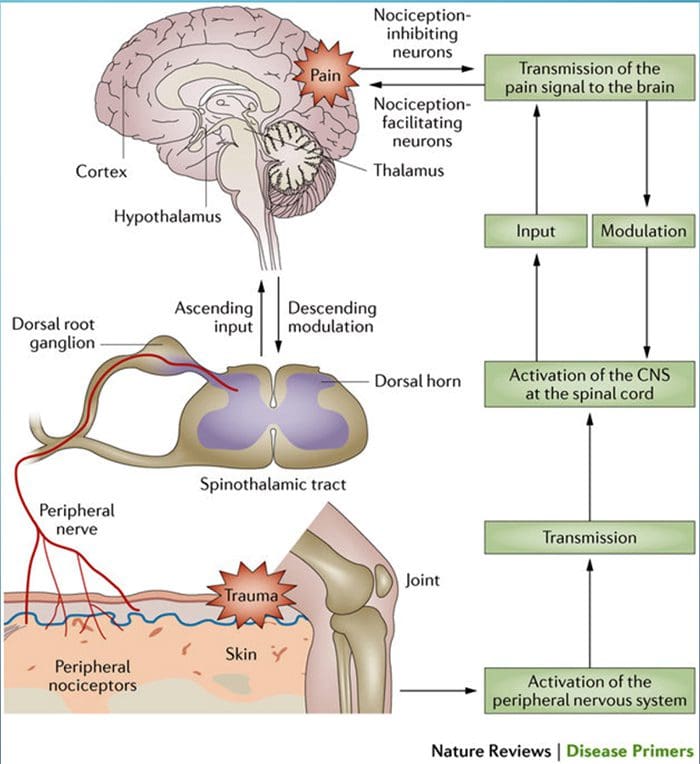
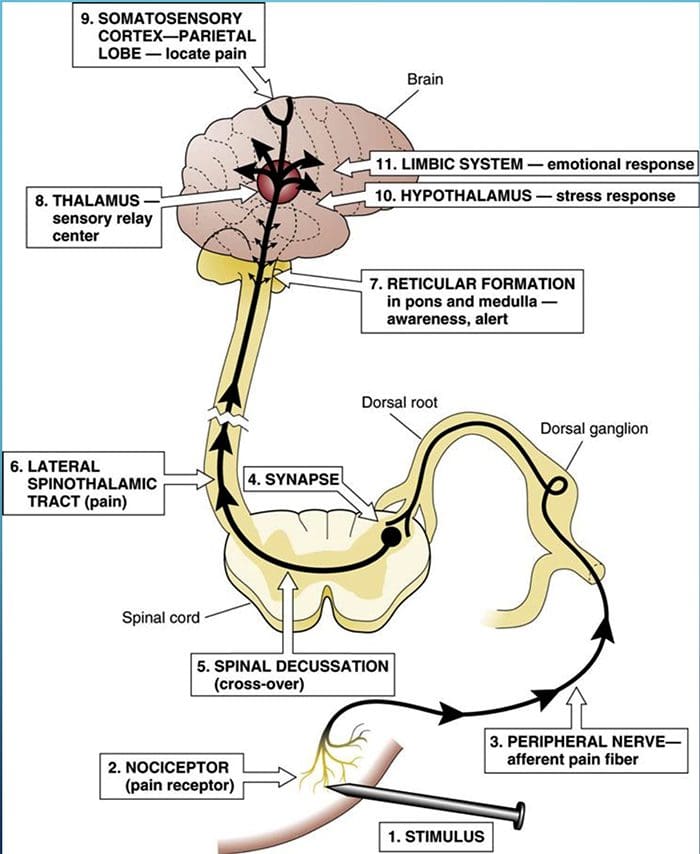
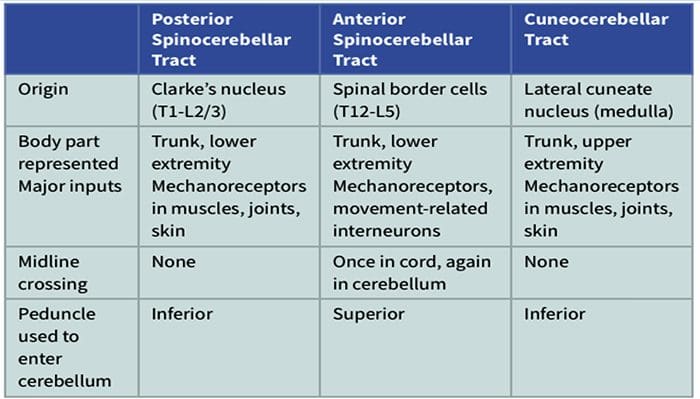
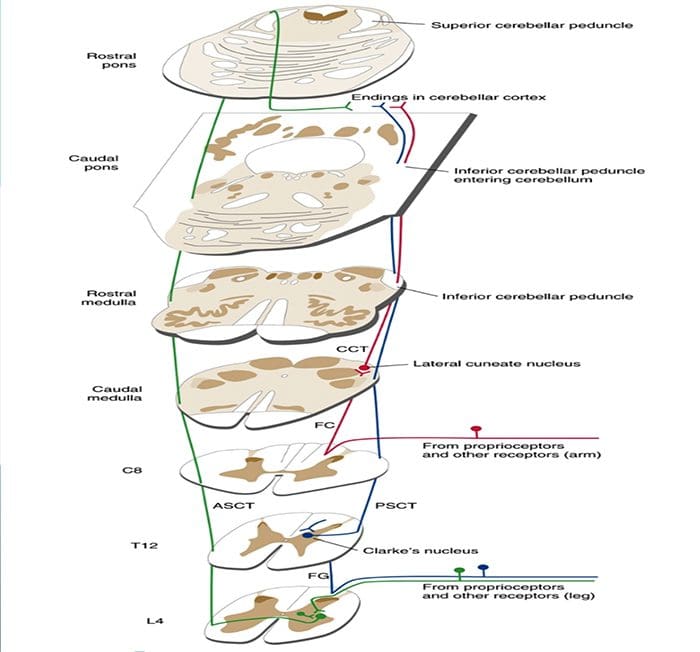
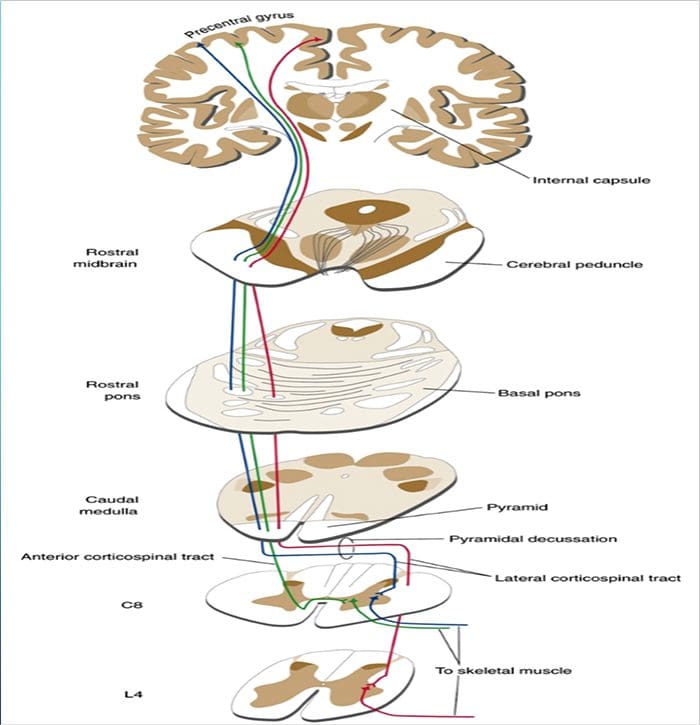
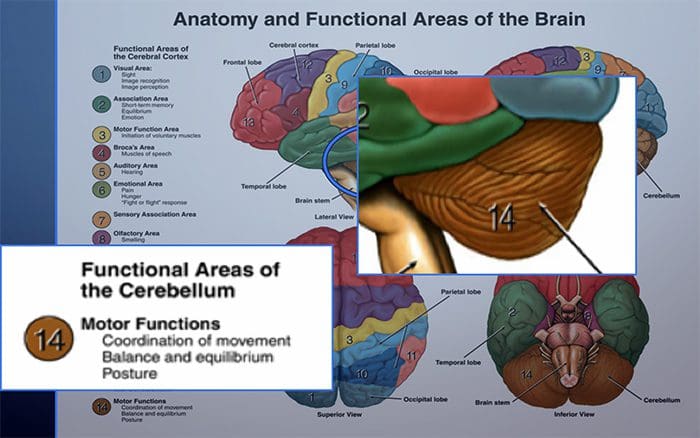
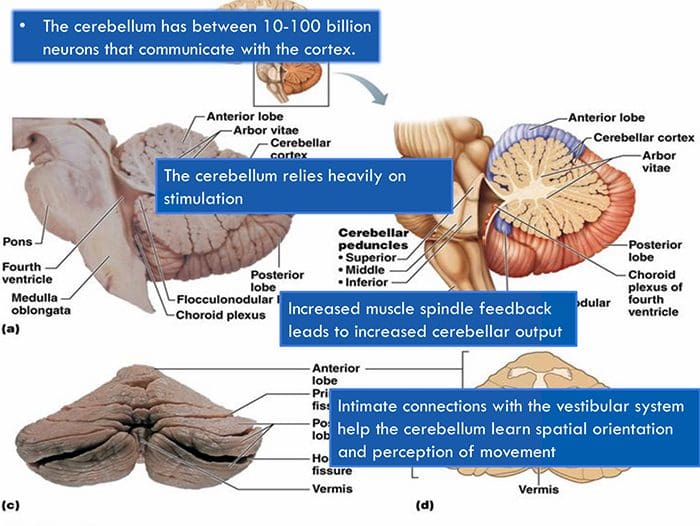
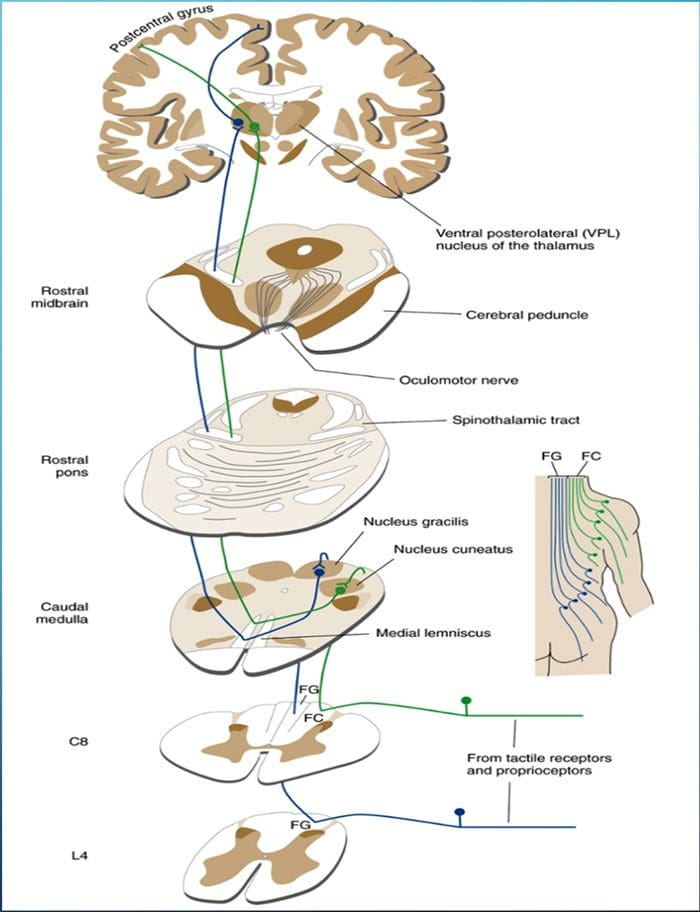 Kolom posterior merujuk pada seluruh isi funiculus posterior, eksklusif bagiannya dari saluran propriospinal. Kolom posterior terutama terdiri dari kolateral naik dari aferen utama myelinated besar membawa impuls dari berbagai macam mechanoreceptors (meskipun sejumlah besar serat orde kedua dan serat unmyelinated juga termasuk). Ini secara tradisional dianggap sebagai jalur utama di mana informasi dari reseptor kulit, sendi, dan otot ambang rendah mencapai korteks serebri.
Kolom posterior merujuk pada seluruh isi funiculus posterior, eksklusif bagiannya dari saluran propriospinal. Kolom posterior terutama terdiri dari kolateral naik dari aferen utama myelinated besar membawa impuls dari berbagai macam mechanoreceptors (meskipun sejumlah besar serat orde kedua dan serat unmyelinated juga termasuk). Ini secara tradisional dianggap sebagai jalur utama di mana informasi dari reseptor kulit, sendi, dan otot ambang rendah mencapai korteks serebri.