
Kami semua diajarkan sebagai anak-anak yang ada indra 5: penglihatan, rasa, suara, bau, dan sentuhan. Empat indra awal menggunakan organ-organ yang jelas dan berbeda, seperti mata, lidah, telinga, dan hidung, tetapi bagaimana sentuhan indra tubuh secara tepat? Sentuhan dialami di seluruh tubuh, baik di dalam maupun di luar. Tidak ada organ yang berbeda yang bertanggung jawab untuk merasakan sentuhan. Sebaliknya, ada reseptor kecil, atau ujung saraf, di sekitar seluruh tubuh yang merasakan sentuhan di mana itu terjadi dan mengirimkan sinyal ke otak dengan informasi mengenai jenis sentuhan yang terjadi. Sebagai pengecap lidah di lidah mendeteksi rasa, mechanoreceptors adalah kelenjar di dalam kulit dan pada organ lain yang mendeteksi sensasi sentuhan. Mereka dikenal sebagai mechanoreceptors karena mereka dirancang untuk mendeteksi sensasi mekanis atau perbedaan tekanan.
Konten
Peran Mechanoreceptors
Seseorang memahami bahwa mereka telah mengalami sensasi setelah organ yang bertanggung jawab untuk menemukan arti tertentu mengirimkan pesan ke otak, yang merupakan organ utama yang memproses dan mengatur semua informasi. Pesan dikirim dari semua area tubuh ke otak melalui kabel yang disebut sebagai neuron. Ada ribuan neuron kecil yang bercabang ke seluruh area tubuh manusia, dan pada ujung dari banyak neuron ini adalah mechanoreceptors. Untuk menunjukkan apa yang terjadi ketika Anda menyentuh suatu objek, kita akan menggunakan contoh.
Membayangkan seekor nyamuk mendarat di lengan Anda. Strain serangga ini, sangat ringan, menstimulasi mechanoreceptors di area tertentu pada lengan. Mekanoreptor tersebut mengirim pesan di sepanjang neuron yang terhubung dengannya. Neuron menghubungkan semua jalan ke otak, yang menerima pesan bahwa ada sesuatu yang menyentuh tubuh Anda di lokasi yang tepat dari mechanoreceptor tertentu yang mengirim pesan. Otak akan bertindak dengan saran ini. Mungkin itu akan memberi tahu mata untuk melihat wilayah lengan yang mendeteksi tanda tangan. Dan ketika mata mengatakan pada otak bahwa ada nyamuk di lengannya, otak mungkin akan memberitahu tangan untuk segera membuangnya. Begitulah cara kerja mechanoreceptors. Tujuan artikel di bawah ini adalah untuk mendemonstrasikan serta mendiskusikan secara detail organisasi fungsional dan determinan molekul dari mechanoreceptors.
Sentuh Sense: Organisasi Fungsional dan Penentu Molekul Penentu Mekanisme
Abstrak
Mechanoreceptors kulit dilokalisasi di berbagai lapisan kulit di mana mereka mendeteksi berbagai rangsangan mekanik, termasuk sikat cahaya, peregangan, getaran dan tekanan berbahaya. Berbagai rangsangan ini dicocokkan oleh beragam mekanisme mekanoreptor khusus yang merespon deformasi kulit dengan cara tertentu dan menyampaikan rangsangan ini ke struktur otak yang lebih tinggi. Studi di seluruh mechanoreceptors dan ujung saraf sensorik traktus mulai menemukan mekanisme sentuhan sensasi. Bekerja di bidang ini telah memberikan para peneliti dengan pemahaman yang lebih menyeluruh dari organisasi sirkuit yang mendasari persepsi sentuhan. Saluran ion baru telah muncul sebagai kandidat untuk molekul transduksi dan sifat arus mekanis secara mekanis meningkatkan pemahaman kita tentang mekanisme adaptasi terhadap rangsangan taktil. Ulasan ini menyoroti kemajuan yang dibuat dalam mengkarakterisasi sifat fungsional dari mechanoreceptors di kulit berbulu dan gabut dan saluran ion yang mendeteksi input mekanik dan membentuk adaptasi mechanoreceptor.
Kata kunci: mechanoreceptor, saluran sensitif mekanis, nyeri, kulit, sistem somatosensori, sentuhan
Pengantar
Sentuhan adalah deteksi rangsangan mekanik yang berdampak pada kulit, termasuk rangsangan mekanik yang tidak berbahaya dan berbahaya. Ini adalah perasaan penting untuk kelangsungan hidup dan perkembangan mamalia dan manusia. Kontak benda padat dan cairan dengan kulit memberikan informasi yang diperlukan ke sistem saraf pusat yang memungkinkan eksplorasi dan pengenalan lingkungan dan memulai pergerakan atau gerakan tangan yang direncanakan. Sentuhan juga sangat penting untuk pemagangan, kontak sosial dan seksualitas. Indera sentuhan adalah indra yang paling tidak rentan, meskipun dapat terdistorsi (hyperesthesia, hypoesthesia) di banyak kondisi patologis.1-3
Respons sentuhan melibatkan pengkodean informasi mekanis yang sangat tepat. Mekanoreseptor kulit terlokalisasi di berbagai lapisan kulit tempat mereka mendeteksi berbagai rangsangan mekanis, termasuk sikat ringan, regangan, getaran, defleksi rambut, dan tekanan berbahaya. Variasi rangsangan ini dicocokkan dengan beragam mekanoreseptor khusus yang merespons deformasi kulit dengan cara tertentu dan menyampaikan rangsangan ini ke struktur otak yang lebih tinggi. Neuron somatosensori pada kulit terbagi dalam dua kelompok: mekanoreseptor ambang rendah (LTMR) yang bereaksi terhadap tekanan jinak dan mekanoreseptor ambang tinggi (HTMR) yang merespons rangsangan mekanis berbahaya. Badan sel LTMR dan HTMR berada dalam ganglia akar dorsal (DRG) dan ganglia sensorik kranial (ganglia trigeminal). Serat saraf yang terkait dengan LTMR dan HTMR diklasifikasikan sebagai serat A? -, A? - atau C berdasarkan kecepatan konduksi potensial aksi mereka. Serat C tidak bermielin dan memiliki kecepatan konduksi paling lambat (~ 2 m / s), sedangkan A? dan A? serat ringan dan sangat mielin, masing-masing menunjukkan kecepatan konduksi menengah (~ 12 m / s) dan cepat (~ 20 m / s). LTMR juga diklasifikasikan sebagai respons yang lambat, atau cepat beradaptasi (SA- dan RA-LTMR) menurut tingkat adaptasinya terhadap stimulus mekanis yang berkelanjutan. Mereka selanjutnya dibedakan oleh organ ujung kulit yang mereka sediakan dan rangsangan yang mereka sukai.
Kemampuan mechanoreceptors untuk mendeteksi isyarat mekanis bergantung pada kehadiran saluran ion mekanotransducer yang dengan cepat mengubah kekuatan mekanik menjadi sinyal listrik dan depolarise bidang reseptif. Depolarisasi lokal ini, yang disebut potensi reseptor, dapat menghasilkan potensial aksi yang menyebar ke sistem saraf pusat. Namun, sifat molekul yang memediasi mekanotransduksi dan adaptasi terhadap kekuatan mekanik tetap tidak jelas.
Dalam ulasan ini, kami memberikan gambaran umum tentang sifat mechanoreceptor mamalia dalam sentuhan tidak berbahaya dan berbahaya pada kulit berbulu dan licin. Kami juga mempertimbangkan pengetahuan terbaru tentang sifat-sifat arus gerbang mekanis dalam upaya untuk menjelaskan mekanisme adaptasi mechanoreceptor. Akhirnya, kami meninjau kemajuan terbaru yang dibuat dalam mengidentifikasi saluran ion dan protein terkait yang bertanggung jawab untuk menghasilkan arus mechano-gated.
Sentuhan Tidak Sehat
Hair Follicle-Associated LTMRs
Folikel rambut mewakili organ penghasil batang rambut yang mendeteksi sentuhan ringan. Serat yang terkait dengan folikel rambut merespon gerakan rambut dan arahnya dengan menembakan kereta potensial aksi pada awal dan penghilangan stimulus. Mereka dengan cepat mengadaptasi reseptor.
Kucing dan kelinci. Pada bulu kucing dan kelinci, folikel rambut dapat dibagi menjadi tiga jenis folikel rambut, yaitu Rambut Bawah, Rambut Penjaga dan Tylotrichs. The Down hairs (underhair, wool, vellus) 4 adalah bulu yang paling banyak, terpendek dan paling halus dari mantel. Mereka bergelombang, tidak berwarna dan muncul dalam kelompok dua hingga empat rambut dari lubang umum di kulit. The Guard hairs (monotrichs, overhears, tophair) 4 sedikit melengkung, baik berpigmen atau tidak berpigmen, dan muncul sendiri-sendiri dari mulut folikel mereka. Tylotrichs adalah yang paling sedikit jumlahnya, rambut terpanjang dan paling tebal.5,6 Mereka berpigmen atau tidak berpigmen, kadang-kadang keduanya dan muncul sendiri-sendiri dari folikel yang dikelilingi oleh lingkaran pembuluh darah kapiler. Pasokan serat sensorik ke folikel rambut terletak di bawah kelenjar sebaceous dan dihubungkan dengan A? atau serat A? -LTMR.7
Dalam aposisi dekat ke batang rambut bawah, tepat di bawah tingkat kelenjar sebaceous adalah cincin ujung pilo-Ruffini lanceolate. Ujung-ujung saraf sensorik ini diposisikan dalam sebuah spiral di sekitar batang rambut di dalam jaringan ikat yang membentuk folikel rambut. Di dalam folikel rambut, ada juga ujung saraf bebas, beberapa di antaranya membentuk mechanoreceptors. Sering, menyentuh sel-sel (lihat kulit berkerut) mengelilingi daerah leher folikel tylotrich.
Sifat ujung saraf mielin pada kulit berbulu kucing dan kelinci telah dieksplorasi secara intensif pada periode 1930-1970 (review dalam Hamann, 1995) .8 Hebatnya, Brown dan Iggo, mempelajari 772 unit dengan serabut saraf aferen bermielin pada saraf saphena dari kucing. dan kelinci, telah mengklasifikasikan respons dalam tiga jenis reseptor yang sesuai dengan pergerakan bulu Bawah (reseptor tipe D), Rambut pelindung (reseptor tipe G) dan rambut Tylotrich (reseptor tipe T) .9 Semua respons serat saraf aferen telah disatukan di reseptor Beradaptasi Cepat tipe I (RA I) oleh oposisi terhadap reseptor Pacinian bernama RA II. Mekanoreseptor RA I mendeteksi kecepatan stimulus mekanis dan memiliki batas yang tajam. Mereka tidak mendeteksi variasi termal. Burgess dkk. juga menggambarkan reseptor medan yang cepat beradaptasi yang merespon secara optimal untuk membelai kulit atau pergerakan beberapa rambut, yang dikaitkan dengan stimulasi ujung pilo-Ruffini. Tidak ada respon folikel rambut yang dikaitkan dengan aktivitas serat C.
Tikus. Pada kulit berbulu punggung tikus, tiga jenis utama folikel rambut telah diuraikan: zigzag (sekitar 72%), awl / auchene (sekitar 23%) dan penjaga atau tylotrich (sekitar 5%) 11-14 Zigzag dan Awl / folikel rambut auchenne menghasilkan batang rambut yang lebih tipis dan pendek dan berhubungan dengan satu kelenjar sebaceous. Rambut penjaga atau tylotrich adalah jenis folikel rambut terpanjang. Mereka dicirikan oleh bohlam rambut besar yang dikaitkan dengan dua kelenjar sebaceous. Rambut penjaga dan rambut awl / auchene disusun dalam pola berulang yang teratur, sedangkan rambut zigzag yang padat mengisi area kulit di sekitar dua jenis folikel rambut yang lebih besar [Gambar. 1 (A1, A2, dan A3)].
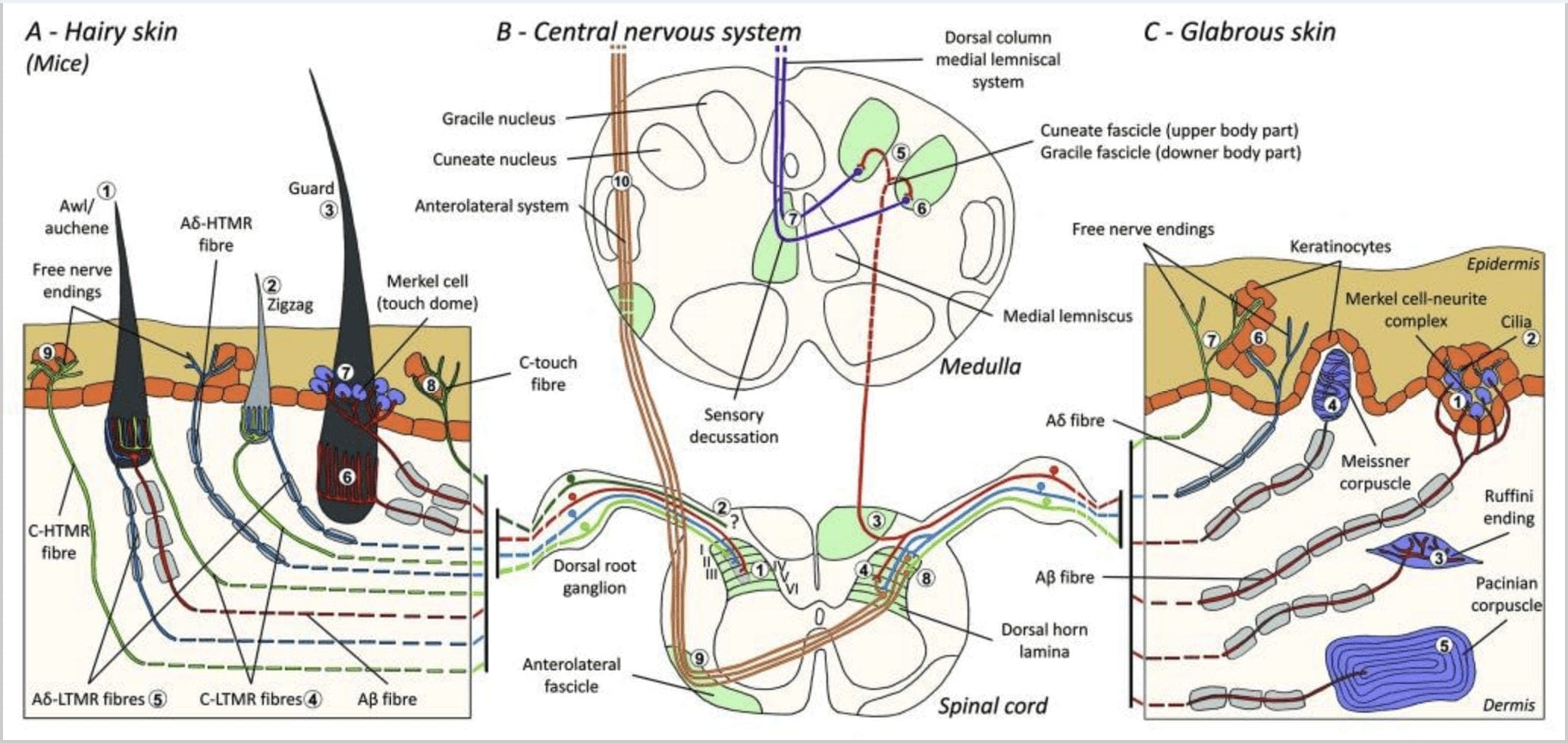
Gambar 1. Organisasi dan proyeksi mekanoreptor kulit. Pada kulit berbulu, sikat dan sentuhan ringan terutama dideteksi oleh persarafan di sekitar folikel rambut: awl / auchenne (A1), zigzag (A2) dan penjaga (A3). Rambut awl / auchene dipersarafi tiga kali oleh ujung lanset C-LTMR (A4), A? -LTMR dan A? cepat beradaptasi-LTMR (A6). Folikel rambut zigzag adalah batang rambut yang lebih pendek dan dipersarafi oleh C-LTMR (A4) dan A? -LTMR ujung lanset (A5). Folikel rambut pelindung terpanjang dipersarafi oleh A? ujung lanset memanjang LTMR beradaptasi dengan cepat (A6) dan berhubungan dengan A? perlahan mengadaptasi-LTMR ujung kubah sentuh (A7). Proyeksi sentral dari semua serat ini berakhir dalam lamina yang berbeda, tetapi sebagian tumpang tindih dari tanduk dorsal sumsum tulang belakang (C-LTMR di lamina II, A? -LTMR di lamina III dan A? -LTMR di lamina IV dan V). Proyeksi dari LTMR yang menginervasi folikel rambut yang sama atau berdekatan selaras untuk membentuk kolom sempit di dorsal dorsal horn tulang belakang (B1 dalam warna abu-abu). Hanya pada kulit berbulu, subpopulasi C-fibers free end innervates epidermis dan merespon sentuhan yang menyenangkan (A8). Serat C-touch ini tidak merespons sentuhan berbahaya dan jalur perjalanannya belum diketahui (B2). Dalam kulit gundul, sentuhan tidak berbahaya dimediasi oleh empat jenis LTMR. Kompleks sel-neurit Merkel ada di lapisan basal epidermis (C1). Mekanoreseptor ini terdiri dari pengaturan antara banyak sel Merkel dan terminal saraf yang membesar dari satu A? serat. Sel Merkel menunjukkan jari seperti proses yang menghubungi keratinosit (C2). Akhir Ruffini terlokalisir di dermis. Ini adalah ujung sensorik encapsulated berbentuk cerutu tipis yang terhubung ke A? serat (C3). Sel darah Meissner terhubung ke A? ujung saraf dan terletak di papila dermal. Mechanoreceptor encapsulated ini terdiri dari dikemas ke bawah sel-sel suportif diatur sebagai lamella horizontal dikelilingi oleh jaringan ikat (C4). Corpus korpus adalah mekanoreptor yang lebih dalam. Satu tunggal A? Ujung saraf tak bermielin berakhir di tengah sel darah bulat telur besar ini yang terbuat dari lamella konsentris. Proyeksi serat A? -LTMR di sumsum tulang belakang dibagi menjadi dua cabang. Cabang pusat utama (B3) naik di sumsum tulang belakang di ipsilateral punggung membentuk cuneate atau gracile fascicles (B5) pada tingkat medulla di mana aferen utama membuat sinaps pertama mereka (B6). Neuron sekunder membuat desus sensorik (B7) untuk membentuk saluran pada lemniscus medial yang naik melalui batang otak ke otak tengah, khususnya di talamus. Cabang kedua dari LTMR berakhir di tanduk dorsal di lamina II, IV, V dan mengganggu transmisi nyeri (B4). Sentuhan berbahaya terdeteksi oleh saraf bebas yang berakhir di epidermis dari keduanya yang berbulu (A9) dan kulit yang gabut (C7). Mekanoreseptor ini adalah akhir dari A? -HTMR dan C-HTMR dalam kontak dekat dengan keratinosit tetangga (C6). A? -HTMR berakhir di lamina I dan V; C-HTMR berakhir di lamina I dan II (B8). Pada tingkat dorsal dorsal spinal cord, aferen primer HTMRs membuat sinaps dengan neuron sekunder yang melintasi garis tengah dan naik ke struktur otak yang lebih tinggi di fikel anterolateral (B9, B10).
Baru-baru ini, Ginty dan kolaborator menggunakan kombinasi pelabelan genetik molekuler dan pendekatan pelacakan retrograde somatotopik untuk memvisualisasikan organisasi ujung aksonal perifer dan sentral dari LTMR pada tikus.15 Temuan mereka mendukung model di mana fitur individu dari stimulus taktil yang kompleks adalah diekstraksi oleh tiga jenis folikel rambut dan disampaikan melalui aktivitas kombinasi unik serat A? -, A? - dan C- ke tanduk punggung.
Mereka menunjukkan bahwa pelabelan genetik dari neuron DRG tirosin hidroksilase positif (TH +) mencirikan populasi neuron sensorik nonpeptidergik, berdiameter kecil dan memungkinkan visualisasi ujung perifer C-LTMR di kulit. Anehnya, cabang aksonal dari C-LTMR individu ditemukan untuk arborise dan membentuk ujung lanset longitudinal yang terkait erat dengan zigzag (80% ujung) dan penusuk / auchene (20% ujung), tetapi tidak folikel rambut tylotrich [Gbr. 1 (A4)]. Ujung lanset memanjang telah lama dianggap milik eksklusif A? -LTMRs dan oleh karena itu tidak terduga bahwa ujung C-LTMR akan membentuk ujung lanset memanjang.15 C-LTMR ini memiliki adaptasi menengah dibandingkan dengan lambat dan cepat beradaptasi. mekanoreseptor bermielin [Gbr. 2 (C1)].
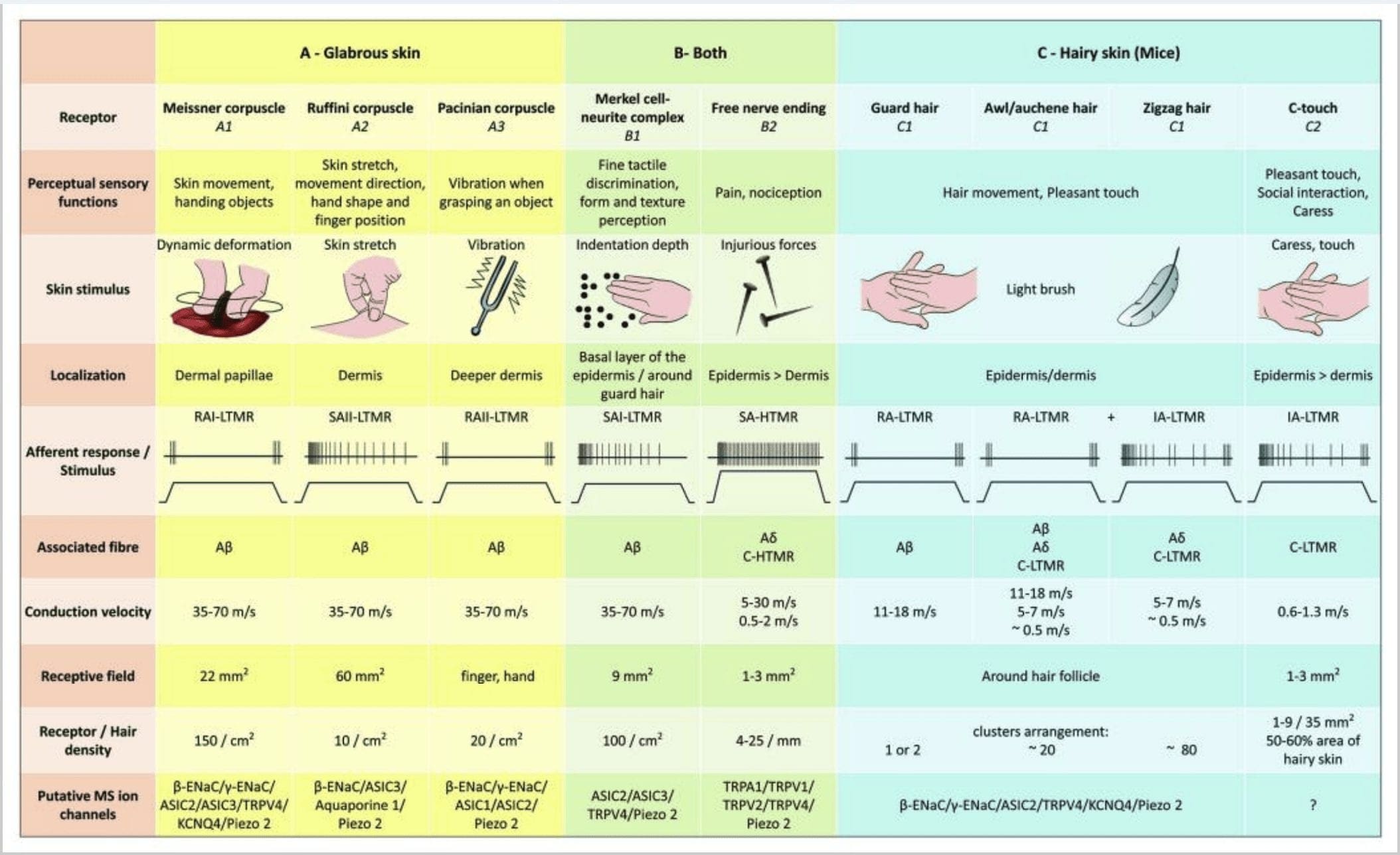
Gambar 2. Reseptor taktil pada mamalia: Reseptor taktil kulit berdiferensiasi menjadi sentuhan tidak berbahaya yang didukung oleh beberapa reseptor dengan ambang mekanis rendah (LTMR) pada kulit gundul dan berbulu serta sentuhan berbahaya yang didukung oleh reseptor ambang mekanis tinggi (HTMRs). Mereka membentuk ujung bebas saraf yang berakhir terutama di epidermis. (A) Kulit berkilau. A1: Sel darah Meissner mendeteksi gerakan kulit dan benda tergelincir di tangan. Mereka penting untuk menyerahkan benda dan ketangkasan. Reseptor cepat beradaptasi dengan stimulus, terhubung ke A? serat dan serat C jarang dan memiliki bidang reseptor yang besar. A2: Ruffini corpuscles mendeteksi peregangan kulit dan penting untuk mendeteksi posisi jari dan benda tangan. Reseptor perlahan beradaptasi dengan rangsangan dan mempertahankan aktivitas selama rangsangan itu diterapkan. Reseptor terhubung ke A? serat dan memiliki bidang reseptif yang besar. A3: Sel-sel Pacinian lebih dalam di dermis dan mendeteksi getaran. Reseptor terhubung ke A? serat; mereka cepat beradaptasi dengan stimulus dan memiliki bidang reseptif terbesar. (B) Seluruh kulit. B1: Kompleks sel merkel ada di kulit yang licin dan di sekitar rambut. Mereka diekspresikan dengan padat di tangan dan penting untuk persepsi tekstur dan diskriminasi terbaik antara dua titik. Mereka bertanggung jawab atas ketepatan jari. Reseptor terhubung ke A? serat; mereka perlahan-lahan beradaptasi dengan stimulus dan memiliki bidang reseptif yang pendek. B2: HTMR sentuhan berbahaya dengan adaptasi yang sangat lambat terhadap stimulus, yaitu aktif selama stimulus nosiseptif diterapkan. Mereka dibentuk oleh ujung saraf bebas A? dan serat-C yang terkait dengan keratinosit. (C) Kulit berbulu. C1: Folikel rambut berhubungan dengan berbagai jenis rambut. Pada tikus, rambut Penjaga adalah yang lebih panjang dan jarang diekspresikan, penusuk / auchenne berukuran sedang dan zigzag adalah rambut terkecil dan paling padat. Mereka terhubung ke A? serat tetapi juga ke A? dan serat C-LTMR untuk rambut awl / auchenne dan zizag. Mereka mendeteksi gerakan rambut termasuk sentuhan yang menyenangkan selama belaian. Mereka beradaptasi dengan cepat atau dengan kinetik menengah terhadap rangsangan. C2: ujung saraf C-touch sesuai dengan subtipe ujung serabut C dengan ujung bebas yang ditandai dengan ambang mekanis rendah. Mereka seharusnya menyandikan untuk sensasi menyenangkan yang disebabkan oleh belaian. Mereka cukup beradaptasi dengan stimulus dan memiliki bidang reseptif yang pendek. Kanal ion mekanosensitif (MS) putatif yang diekspresikan dalam reseptor taktil yang berbeda ditunjukkan sesuai dengan data awal dan meringkas hipotesis saat ini yang sedang dievaluasi.
Populasi besar kedua yang diidentifikasi menyangkut ujung A? -LTMR di folikel Awl / Auchenne dan zigzag untuk dibandingkan dengan folikel bulu Down yang dipelajari secara ekstensif pada kucing dan kelinci. Ginty dan kolaborator menunjukkan bahwa TrkB diekspresikan pada tingkat tinggi dalam subset neuron DRG berdiameter sedang. Rekaman intraseluler menggunakan preparasi saraf kulit ex vivo dari serat berlabel mengungkapkan bahwa mereka menunjukkan sifat fisiologis serat yang sebelumnya dipelajari pada kucing dan kelinci: sensitivitas mekanis yang sangat baik (ambang Von Frey <0.07 mN), respons yang cepat beradaptasi terhadap rangsangan ambang batas, konduksi menengah kecepatan (5.8 0.9 m / s) dan lonjakan sempit soma yang tidak terinfeksi.15 A? -LTMRs ini membentuk ujung lanset longitudinal yang terkait dengan hampir setiap folikel rambut zigzag dan penusuk / auchene dari batang [Gbr. 1 (A5)].
Akhirnya, mereka menunjukkan bahwa ujung perifer dari A? LTMR membentuk ujung lanset longitudinal yang terkait dengan folikel rambut pelindung (atau tylotrich) dan penusuk / auchene [Gambar. 1 (A6)]. 15 Selain itu, bulu pelindung juga dikaitkan dengan kompleks sel Merkel yang membentuk kubah sentuh yang terhubung ke A? perlahan mengadaptasi LTMR [Gbr. 1 (A7)].
Singkatnya, hampir semua folikel rambut zigzag dipersarafi oleh ujung lanset C-LTMR dan A? -LTMR; rambut awl / auchene dipersarafi tiga kali lipat oleh A? ujung lanset yang beradaptasi dengan cepat-LTMR, A? -LTMR dan C-LTMR; Folikel rambut pelindung dipersarafi oleh A? ujung lanset memanjang LTMR beradaptasi dengan cepat dan berinteraksi dengan A? perlahan mengadaptasi-LTMR ujung kubah sentuh. Dengan demikian, setiap folikel rambut tikus menerima kombinasi ujung LTMR yang unik dan tidak berubah sesuai dengan organ ujung mekanosensori yang berbeda secara neurofisiologis. Mempertimbangkan pengaturan berulang dari ketiga jenis rambut ini, Ginty dan kolaborator mengusulkan bahwa kulit berbulu terdiri dari pengulangan berulang unit perifer yang mengandung, (1) satu atau dua rambut pelindung yang terletak di tengah, (2) ~ 20 rambut penusuk / auchenne sekitarnya dan (3 ) ~ 80 rambut zigzag diselingi [Gbr. 2 (C1)].
Proyeksi sumsum tulang belakang. Proyeksi sentral A? LTMR yang cepat beradaptasi, A? -LTMRs dan C-LTMRs berakhir di lamina yang berbeda, tetapi sebagian tumpang tindih (II, III, IV) dari tanduk punggung medula spinalis. Selain itu, terminal pusat LTMR yang menginervasi folikel rambut yang sama atau berdekatan dalam unit LTMR perifer diselaraskan untuk membentuk kolom LTMR sempit di tanduk dorsal sumsum tulang belakang [Gbr. 1 (B1)]. Dengan demikian, nampaknya irisan, atau kolom ujung aferen sensorik primer yang tersusun secara somatotopic di tanduk dorsal mewakili kesejajaran proyeksi pusat A? -, A? - dan C-LTMR yang menginervasi unit perifer yang sama dan mendeteksi rangsangan yang bekerja pada kelompok kecil folikel rambut yang sama. Berdasarkan jumlah bulu pelindung, penusuk / auchene, dan zigzag pada batang dan tungkai serta jumlah setiap subtipe LTMR, Ginty dan kolaborator memperkirakan bahwa cula punggung tikus mengandung 2,000-4,000 kolom LTMR, yang sesuai dengan perkiraan jumlah periferal. Unit LTMR. 15
Lebih lanjut, akson dari subtipe LTMR terkait erat satu sama lain, memiliki proyeksi yang terjalin dan ujung lanset interdigitasi yang menginervasi folikel rambut yang sama. Selain itu, karena ketiga jenis folikel rambut menunjukkan bentuk, ukuran, dan komposisi seluler yang berbeda, mereka cenderung memiliki sifat penyetelan defleksi atau getaran yang berbeda. Temuan ini konsisten dengan pengukuran neurofisiologis klasik pada kucing dan kelinci yang menunjukkan bahwa A? RA-LTMRs dan A? -LTMRs dapat diaktifkan secara berbeda dengan defleksi jenis folikel rambut yang berbeda.
Sebagai kesimpulan, sentuhan pada kulit berbulu adalah kombinasi dari: (1) jumlah relatif, distribusi spasial unik dan sifat morfologi dan defleksi yang berbeda dari ketiga jenis folikel rambut; (2) kombinasi unik akhir subtipe LTMR yang terkait dengan masing-masing dari tiga jenis folikel rambut; dan (3) kepekaan yang berbeda, kecepatan konduksi, pola kereta lonjakan dan sifat adaptasi dari empat kelas utama LTMRs terkait-folikel rambut yang memungkinkan sistem skinosensori kulit berbulu untuk mengekstrak dan menyampaikan kepada CNS kombinasi kompleks kualitas yang menentukan menyentuh.
Ujung Lepas Gratis-Saraf LTMR
Umumnya, ujung bebas serat-C di kulit adalah HTMR, tetapi subpopulasi serat-C tidak merespons sentuhan berbahaya. Bagian dari aferen C-fiber (CT) taktil ini mewakili tipe berbeda dari unit mechanoreceptive tanpa mielin, ambang batas rendah yang ada di kulit manusia dan mamalia yang berbulu tetapi tidak gundul [Gbr. 1 (A8)]. 18,19 CT umumnya dikaitkan dengan persepsi stimulasi sentuhan yang menyenangkan dalam kontak tubuh.
Aferen CT merespons gaya lekukan dalam kisaran 0.3 2.5 mN dan karenanya sensitif terhadap deformasi kulit sebanyak A? aferen.19 Karenanya, karakteristik adaptasi aferen CT adalah perantara dibandingkan dengan mekanoreseptor mielin yang beradaptasi dengan lambat dan cepat. Bidang reseptif aferen CT manusia berbentuk bulat atau oval. Bidang ini terdiri dari satu hingga sembilan titik responsif kecil yang tersebar di area hingga 35 mm2.22 Reseptor homolog tikus diatur dalam pola tambalan terputus-putus yang menutupi sekitar 50-60% area di kulit berbulu [Gbr. 2 (C2)]. 23
Bukti dari pasien yang kekurangan afferen sentuhan myelinated menunjukkan bahwa sinyal pada serat CT mengaktifkan korteks insular. Karena sistem ini buruk dalam menyandikan aspek-aspek diskriminatif dari sentuhan, tetapi sangat cocok untuk menyandikan lambat, sentuhan lembut, serat CT pada kulit berbulu dapat menjadi bagian dari sistem untuk memproses aspek sentuhan yang menyenangkan dan relevan secara sosial. Aktivasi serat CT 24 juga mungkin memiliki peran dalam penghambatan rasa sakit dan baru-baru ini telah diusulkan bahwa peradangan atau trauma dapat mengubah sensasi yang disampaikan oleh C-fiber LTMRs dari sentuhan yang menyenangkan ke rasa sakit.25,26
Perjalanan CT-aferen jalur mana yang belum diketahui [Gambar. 1 (B2)], tetapi input taktil threshold rendah ke sel-sel proyeksi spinothalamik telah didokumentasikan, 27 meminjamkan kepercayaan pada laporan-laporan defisit deficit yang halus dan kontralateral pada pasien manusia setelah penghancuran jalur ini setelah prosedur chordotomy.28
LTMRs di Glabrous Skin
Kompleks sel-neurite merkel dan kubah sentuh. Merkel (1875) adalah orang pertama yang memberikan deskripsi histologis kelompok sel epidermis dengan inti berlobus besar, melakukan kontak dengan serabut saraf aferen yang diduga. Dia berasumsi bahwa mereka mengabdi pada indra peraba dengan menyebut mereka Tastzellen (sel taktil). Pada manusia, kompleks neurit sel Merkel diperkaya pada area sensitif sentuhan pada kulit, ditemukan di lapisan basal epidermis di jari, bibir dan alat kelamin. Mereka juga ada di kulit berbulu dengan kepadatan lebih rendah. Kompleks neurit sel Merkel terdiri dari sel Merkel yang terletak dekat dengan terminal saraf yang membesar dari A? serat [Gambar. 1 (C1)] (review di Halata dan kolaborator) .29 Di sisi epidermal sel Merkel menunjukkan proses seperti jari yang membentang di antara keratinosit tetangga [Gambar. 1 (C2)]. Sel Merkel adalah sel epidermis turunan keratinosit. 30,31 Istilah kubah sentuh diperkenalkan untuk menamai konsentrasi besar kompleks sel Merkel pada kulit berbulu kaki depan kucing. Kubah sentuh dapat memiliki hingga 150 sel Merkel yang diinervasi oleh satu serat A? Dan pada manusia selain A? -Fibers, A? dan C-fiber juga sering ditemukan. 32-34
Stimulasi kompleks neurit sel Merkel menghasilkan respons Tipe I (SA I) yang beradaptasi perlahan, yang berasal dari bidang reseptif tanda baca dengan batas tajam. Tidak ada pelepasan spontan. Kompleks ini merespons kedalaman lekukan kulit dan memiliki resolusi spasial tertinggi (0.5 mm) dari mekanoreseptor kulit. Mereka mengirimkan gambar spasial yang tepat dari rangsangan taktil dan diusulkan untuk bertanggung jawab atas diskriminasi bentuk dan tekstur [Gbr. 2 (B1)]. Tikus tanpa sel Merkel tidak dapat mendeteksi permukaan bertekstur dengan kakinya saat mereka melakukannya menggunakan kumis
Apakah sel Merkel, neuron sensorik atau keduanya merupakan tempat mekanotransduksi masih menjadi bahan perdebatan. Pada tikus, penghancuran fototoksik sel Merkel menghilangkan respons SA I. Pada tikus dengan sel Merkel yang tertekan secara genetik, respons SA I yang direkam dalam preparasi kulit / saraf ex vivo benar-benar menghilang, menunjukkan bahwa sel Merkel diperlukan untuk pengkodean Merkel yang tepat. respon reseptor.36 Namun, stimulasi mekanis sel Merkel terisolasi dalam kultur oleh tekanan yang digerakkan motor tidak menghasilkan arus gerbang mekanis.37 Keratinosit mungkin memainkan peran penting dalam fungsi normal kompleks neurit sel Merkel. Proses seperti jari sel Merkel dapat bergerak dengan deformasi kulit dan pergerakan sel epidermis, dan ini mungkin merupakan langkah pertama dari transduksi mekanis. Jelas, kondisi yang diperlukan untuk mempelajari sensitivitas mechano sel Merkel belum ditetapkan.
Akhir Ruffini. Ujung ruffini adalah ujung sensorik encapsulated tipis berbentuk cerutu yang terhubung ke A? ujung saraf. Ujung ruffini adalah silinder jaringan ikat kecil yang tersusun sepanjang untaian kolagen dermal yang disuplai oleh satu hingga tiga serabut saraf bermielin dengan diameter 4 6 m. Hingga tiga silinder dengan orientasi berbeda di dermis dapat bergabung untuk membentuk satu reseptor [Gbr. 1 (C3)]. Secara struktural, ujung Ruffini mirip dengan organ tendon Golgi. Mereka diekspresikan secara luas dalam dermis dan telah diidentifikasi sebagai mekanoreseptor kulit tipe II (SA II) yang beradaptasi perlahan. Dengan latar belakang aktivitas saraf spontan, pelepasan reguler yang beradaptasi secara perlahan ditimbulkan oleh stimulasi mekanis yang dipertahankan gaya rendah tegak lurus atau lebih efektif dengan peregangan kulit. Respons SA II berasal dari bidang reseptif yang besar dengan batas yang tidak jelas. Reseptor Ruffini berkontribusi pada persepsi arah gerak benda melalui pola peregangan kulit [Gbr. 2 (A2)].
Pada tikus, respon SA I dan SA II dapat dipisahkan secara elektrofisiologi dalam persiapan saraf-kulit ex-vivo. 40 Nandasena dan kolaborator melaporkan imunolocisasi aquaporin 1 (AQP1) pada ujung Ruffini periodontal pada insisivus tikus yang menunjukkan bahwa AQP1 terlibat dalam pemeliharaan keseimbangan osmotik gigi yang diperlukan untuk mekanotransduksi.41 Ujung Ruffini periodontal juga menyatakan saluran ion mechanosensitive diduga ASIC3.42
Meissner corpuscles. Sel-sel Meissner terlokalisasi di papila dermal kulit gundul, terutama di telapak tangan dan telapak kaki, tetapi juga di bibir, di lidah, di wajah, di puting dan di alat kelamin. Secara anatomis, mereka terdiri dari ujung saraf yang dienkapsulasi, kapsul terbuat dari sel pendukung pipih yang disusun sebagai lamella horizontal yang tertanam dalam jaringan ikat. Ada satu serabut saraf A? aferen terhubung per sel [Gbr. 1 (C4)]. Setiap deformasi fisik sel darah memicu serangkaian potensial aksi yang dengan cepat berhenti, yaitu, mereka dengan cepat mengadaptasi reseptor. Ketika rangsangan dihilangkan, sel darah mendapatkan kembali bentuknya dan saat melakukannya menghasilkan potensi aksi lainnya. Karena lokasinya yang dangkal di dermis, sel-sel ini secara selektif merespons gerakan kulit, deteksi sentuhan slip dan getaran (20-40 Hz). Mereka sensitif terhadap kulit dinamis - misalnya, antara kulit dan benda yang sedang ditangani [Gbr. 2 (A1)].
Korpuscles Pacinian. Sel-sel Pacinian adalah mechanoreceptors kulit yang lebih dalam dan merupakan mechanoreceptor kulit terenkapsulasi yang paling sensitif dari gerakan kulit. Sel-sel besar berbentuk bulat telur ini (panjangnya 1 mm) terbuat dari lamellae konsentris dari jaringan ikat fibrosa dan fibroblas yang dilapisi oleh sel Schwann yang dimodifikasi datar diekspresikan di dalam dermis.43 Di tengah sel, dalam rongga berisi cairan yang disebut bola dalam , mengakhiri satu A tunggal? ujung saraf aferen unmyelinated [Gbr. 1 (C5)]. Mereka memiliki bidang reseptif yang besar di permukaan kulit dengan pusat yang sangat sensitif. Perkembangan dan fungsi beberapa tipe mechanoreceptor yang beradaptasi dengan cepat terganggu pada tikus mutan c-Maf. Secara khusus, sel-sel Pacinian mengalami atrofi yang parah
Sel darah pacinian menunjukkan adaptasi yang sangat cepat sebagai respons terhadap lekukan kulit, pelepasan saraf II (RA II) yang beradaptasi dengan cepat yang mampu mengikuti rangsangan getaran frekuensi tinggi, dan memungkinkan persepsi peristiwa jauh melalui getaran yang ditransmisikan. aferen merespons lekukan berkelanjutan dengan aktivitas transien di awal dan mengimbangi stimulus. Mereka juga disebut detektor percepatan karena mereka dapat mendeteksi perubahan dalam kekuatan stimulus dan, jika laju perubahan dalam stimulus diubah (seperti yang terjadi dalam getaran), responsnya menjadi proporsional dengan perubahan ini. Sel-sel Pacinian merasakan perubahan tekanan besar dan sebagian besar dari semua getaran (45 150 Hz), yang dapat dideteksi bahkan sejauh beberapa sentimeter [Gbr. 300 (A2)].
Respon tonik diamati pada korpuscle Pacinian decapsulated.46 Selain itu, korpuscles Pacinian utuh merespon dengan aktivitas berkelanjutan selama rangsangan indentasi konstan, tanpa mengubah ambang mekanik atau frekuensi respon ketika pensinyalan yang dimediasi GABA diblokir antara glia pipih dan akhiran saraf.47 Dengan demikian, komponen non-saraf dari korpuscle Pacinian mungkin memiliki peran ganda dalam menyaring stimulus mekanik serta dalam modulasi sifat respon dari neuron sensorik.
Proyeksi sumsum tulang belakang. Proyeksi A? -LTMR di sumsum tulang belakang dibagi menjadi dua cabang. Cabang sentral utama naik di sumsum tulang belakang di kolom punggung ipsilateral ke tingkat serviks [Gambar. 1 (B3)]. Cabang sekunder berakhir di tanduk dorsal di lamina IV dan mengganggu transmisi nyeri, misalnya. Ini dapat mengurangi rasa sakit sebagai bagian dari kontrol gerbang [Gbr. 1 (B4)]. 48
Pada tingkat serviks, axones dari cabang utama terpisah dalam dua saluran: saluran midline terdiri dari fascicle gracile yang menyampaikan informasi dari bagian bawah tubuh (kaki dan batang), dan saluran luar terdiri dari rangsangan cuneate yang menyampaikan informasi dari setengah bagian atas. dari tubuh (lengan dan batang) [Gambar. 1 (B5)].
Aferen taktil primer membuat sinaps pertama mereka dengan neuron orde dua di medula di mana serat dari setiap saluran bersinaps di nukleus dengan nama yang sama: akson fasikulus gracile bersinaps di nukleus gracile dan akson cuneate sinaps di nukleus cuneate [Gbr. 1 (B6)]. Neuron yang menerima sinaps menyediakan aferen sekunder dan segera melintasi garis tengah untuk membentuk saluran di sisi kontralateral batang otak - lemniskus medial - yang naik melalui batang otak ke stasiun relai berikutnya di otak tengah, khususnya di talamus [Gambar . 1 (B7)].
Spesifikasi molekuler dari LTMR. Mekanisme molekuler yang mengendalikan diversifikasi awal LTMR baru-baru ini sebagian telah dijelaskan. Bourane dan kolaborator telah menunjukkan bahwa populasi neuron mengekspresikan reseptor Ret tirosin kinase (Ret) dan rekan reseptor GFR? 2 pada tikus embrionik E11 DRG secara selektif mengekspresikan faktor transkripsi Mafa.13 Para penulis ini menunjukkan bahwa Mafa / Neuron Ret / GFR? 49,50 yang ditakdirkan menjadi tiga jenis LTRM spesifik saat lahir: neuron SA2 yang menginervasi kompleks sel Merkel, neuron yang beradaptasi cepat yang menginervasi sel Meissner, dan aferen cepat beradaptasi (RA I) yang membentuk ujung lanset di sekitar folikel rambut. Ginty dan kolaborator juga melaporkan bahwa neuron DRG yang mengekspresikan Ret awal dengan cepat mengadaptasi mechanoreceptors dari sel-sel Meissner, sel-sel Pacinian, dan ujung lanset di sekitar folikel rambut.1 Mereka menginervasi zona target terpisah di dalam nukleus gracile dan cuneate, mengungkapkan pola mekanosensorik modalitas tertentu. proyeksi aksonal neuron dalam batang otak.
Eksplorasi mekanoreptor kulit manusia. Teknik mikroneurografi yang dijelaskan oleh Hagbarth dan Vallbo pada tahun 1968 telah diterapkan untuk mempelajari perilaku pelepasan ujung mekanik tunggal manusia yang memasok otot, sendi dan kulit (lihat untuk review Macefield, 2005) .52,53 Mayoritas mikroneurografi kulit manusia penelitian telah mengkarakterisasi fisiologi aferen taktil pada kulit tangan yang licin. Rekaman mikroelektroda dari median dan saraf ulnaris pada subjek manusia telah mengungkapkan sensasi sentuhan yang dihasilkan oleh empat kelas LTMR: aferen Meissner sangat sensitif terhadap sapuan cahaya di kulit, merespons gaya geser lokal dan slip yang baru jadi atau nyata dalam bidang reseptif. Aferen Pacinian sangat sensitif terhadap transien mekanis cepat. Aferen menanggapi dengan penuh semangat untuk melewati bidang reseptif. Sel darah Pacinian yang terletak di digit biasanya akan merespons ketukan meja yang menopang lengan. Aferen Merkel secara khas memiliki sensitivitas dinamis yang tinggi terhadap rangsangan lekukan yang diterapkan ke area diskrit dan sering merespons dengan pelepasan selama pelepasan. Meskipun aferen Ruffini merespons gaya yang diterapkan secara normal pada kulit, fitur unik aferen SA II adalah kemampuannya untuk merespons juga peregangan kulit lateral. Akhirnya, unit rambut di lengan bawah memiliki bidang ovoid besar atau bidang reseptif tidak beraturan yang terdiri dari beberapa titik sensitif yang berhubungan dengan rambut individu (masing-masing suplai aferen ~ 20 rambut).
Sensitivitas mekanik Keratinosit
Setiap rangsangan mekanik pada kulit harus ditularkan melalui keratinosit yang membentuk epidermis. Sel-sel di mana-mana ini dapat melakukan fungsi pemberian sinyal di samping peran pendukung atau pelindung mereka. Misalnya, keratinosit mensekresi ATP, molekul pemberi sinyal penting, sebagai respons terhadap rangsangan mekanik dan osmotik.54,55 Pelepasan ATP menginduksi peningkatan kalsium intraseluler oleh stimulasi autokrin reseptor purinergik.55 Selanjutnya, ada bukti bahwa hipotonitas mengaktifkan Rho-kinase jalur pensinyalan dan pembentukan serat tegangan F-aktin berikutnya menunjukkan bahwa deformasi mekanik keratinosit dapat mengganggu secara mekanis sel tetangga seperti sel Merkel untuk sentuhan tidak berbahaya dan ujung bebas serat C untuk sentuhan berbahaya [Gambar. 1 (C6)] .56,57
Sentuhan Berbahaya
Mekanoreseptor ambang batas tinggi (HTMR) adalah epidermal C- dan A? ujung saraf bebas. Mereka tidak terkait dengan struktur khusus dan diamati pada kedua kulit berbulu [Gbr. 1 (A9)] dan kulit licin [Gbr. 1 (C7)]. Namun, istilah ujung saraf bebas harus dipertimbangkan dengan hati-hati karena ujung saraf selalu berdekatan dengan keratinosit atau sel Langherans atau melanosit. Analisis ultrastruktural dari ujung saraf mengungkapkan adanya retikulum endoplasma kasar, mitokondria yang melimpah, dan vesikel inti padat. Membran sel epidermis yang berdekatan menebal dan menyerupai membran pasca-sinaptik di jaringan saraf. Perhatikan bahwa interaksi antara ujung saraf dan sel epidermis mungkin dua arah karena sel epidermis dapat melepaskan mediator sebagai ATP, interleukin (IL6, IL10) dan bradikinin dan sebaliknya ujung saraf peptidergik dapat melepaskan peptida seperti CGRP atau substansi P yang bekerja pada sel epidermis. HTMRs terdiri dari mechano-nociceptors yang dieksitasi hanya oleh rangsangan mekanik berbahaya dan nociceptors polimodal yang juga merespon panas berbahaya dan bahan kimia eksogen [Gambar. 2 (B2)]. 58
Serabut aferen HTMR berhenti pada neuron proyeksi di tanduk dorsal medula spinalis. A? -HTMRs menghubungi neuron urutan kedua terutama di lamina I dan V, sedangkan C-HTMR berakhir di lamina II [Gambar. 1 (B8)]. Neuron nosiseptif orde kedua memproyeksikan ke sisi controlateral dari sumsum tulang belakang dan naik ke materi putih, membentuk sistem anterolateral. Neuron-neuron ini berhenti terutama di talamus [Gambar. 1 (B9 dan B10)].
Mechano-Arus dalam Somatosensori Neurones
Mekanisme adaptasi lambat atau cepat dari mechanoreceptors belum dijelaskan. Tidak jelas sejauh mana adaptasi mechanoreceptor disediakan oleh lingkungan seluler dari ujung saraf sensorik, sifat intrinsik dari saluran mekanis-gated dan sifat dari saluran ion tegangan-gated ion di neuron sensorik (Gambar 2). Namun, kemajuan terbaru dalam karakterisasi arus mekanis-gated telah menunjukkan bahwa berbagai kelas saluran mechanosensitive ada di neuron DRG dan dapat menjelaskan beberapa aspek dari adaptasi mechanoreceptors.
Rekaman in vitro pada hewan pengerat telah menunjukkan bahwa soma neuron DRG secara intrinsik bersifat mekanosensitif dan mengekspresikan arus mechano-gated kationik.59-64 Gadolinium dan ruthenium red sepenuhnya memblokir arus mekanosensitif, sedangkan kalsium dan magnesium eksternal, pada konsentrasi fisiologis, serta amilorida dan benzamil, menyebabkan blok parsial.60,62,63 FM1-43 bertindak sebagai penghambat yang bertahan lama, dan injeksi FM1-43 ke kaki belakang tikus menurunkan sensitivitas nyeri pada uji Randall Selitto dan meningkatkan ambang penarikan kaki yang dinilai dengan rambut von Frey.65
Menanggapi stimulasi mekanis yang berkelanjutan, arus mekanosensitif menurun melalui penutupan. Berdasarkan konstanta waktu peluruhan arus, empat jenis arus mechanosensitive yang berbeda telah dibedakan: arus yang beradaptasi dengan cepat (~ 3 6 ms), arus yang beradaptasi secara menengah (~ 15 30 ms), arus yang beradaptasi perlahan (~ 200 300 ms) ) dan arus adaptasi yang sangat lambat (~ 1000 ms) .64 Semua arus ini hadir dengan insiden variabel pada neuron DRG tikus yang menginervasi kulit kaki belakang yang licin.64
Sensitivitas mekanis arus mechanosensitive dapat ditentukan dengan menerapkan serangkaian rangsangan mekanis tambahan, memungkinkan analisis arus stimulus yang relatif rinci.66 Hubungan arus stimulus biasanya sigmoidal, dan amplitudo maksimum arus ditentukan oleh jumlah saluran yang secara bersamaan terbuka.64,67 Menariknya, arus sensitif mekanis yang beradaptasi dengan cepat telah dilaporkan menampilkan ambang mekanis rendah dan titik tengah setengah aktivasi dibandingkan dengan arus sensitif mekanis yang sangat lambat.
Neuron sensorik dengan fenotip non-nociceptive lebih suka mengekspresikan secara cepat mengadaptasikan arus mekanositif dengan ambang mekanis yang lebih rendah.60,61,63,64,68 Sebaliknya, perlahan dan sangat lambat mengadaptasi arus sensitif mekanis kadang-kadang dilaporkan pada sel non-nociceptive yang diduga. 64,68 Ini mendorong saran bahwa arus ini mungkin berkontribusi pada ambang mekanik yang berbeda terlihat pada LTMR dan HTMR in vivo. Meskipun percobaan in vitro ini harus dilakukan dengan hati-hati, dukungan untuk kehadiran di soma dari neuron DRG dari mekanotransduser rendah dan tinggi ambang juga disediakan oleh stimulasi berbasis peregangan radial dari neuron sensorik tikus berbudaya. 69 Paradigma ini mengungkapkan dua populasi utama neuron peregangan sensitif, yang merespon amplitudo stimulus rendah dan satu lagi yang secara selektif merespon amplitudo stimulus tinggi.
Hasil ini memiliki implikasi mekanistik yang penting, namun spekulatif: ambang mekanik neuron sensorik mungkin tidak ada hubungannya dengan organisasi seluler dari mechanoreceptor tetapi mungkin terletak pada sifat-sifat saluran ion mekanis-gated.
Mekanisme yang mendasari desensitisasi arus kation mechanosensitive dalam neuron DRG tikus baru-baru ini telah diurai.64,67 Ini hasil dari dua mekanisme bersamaan yang memengaruhi properti saluran: adaptasi dan inaktivasi. Adaptasi pertama kali dilaporkan dalam penelitian sel rambut pendengaran. Hal ini dapat dijelaskan secara operasional sebagai terjemahan sederhana dari kurva aktivasi saluran transduser sepanjang sumbu stimulus mekanis.70-72 Adaptasi memungkinkan reseptor sensorik untuk mempertahankan kepekaannya terhadap rangsangan baru dengan adanya stimulus yang ada. Namun, sebagian besar arus mekanosensitif dalam neuron DRG tidak dapat diaktifkan kembali setelah stimulasi mekanis pengkondisian, yang menunjukkan inaktivasi beberapa saluran transduser.64,67 Oleh karena itu, inaktivasi dan adaptasi bertindak bersama-sama untuk mengatur arus mekanosensitif. Kedua mekanisme ini umum untuk semua arus mechanosensitive yang diidentifikasi dalam neuron DRG tikus, menunjukkan bahwa elemen fisikokimia terkait menentukan kinetika saluran ini.64
Kesimpulannya, menentukan sifat arus mekanositif endogen secara in vitro sangat penting dalam upaya untuk mengidentifikasi mekanisme transduksi pada tingkat molekuler. Variabilitas yang diamati dalam ambang mekanis dan kinetika adaptasi dari arus gerbang mekanis yang berbeda dalam neuron DRG menunjukkan bahwa sifat intrinsik saluran ion dapat menjelaskan, setidaknya sebagian, ambang mekanis dan kinetika adaptasi dari mechanoreceptors yang dijelaskan dalam dekade 1960 80 menggunakan preparat ex vivo.
Protein yang Hemat Energi
Arus ion mekanositif pada neuron somatosensori dikarakterisasi dengan baik, sebaliknya, hanya sedikit yang diketahui tentang identitas molekul yang memediasi mekanotransduksi pada mamalia. Layar genetik di Drosophila dan C. elegans telah mengidentifikasi molekul-molekul mekanotransduksi kandidat, termasuk TRP dan degenerin / epitelial saluran Na + (Deg / ENaC) family. 73 Upaya terbaru untuk menjelaskan dasar molekuler mekanotransduksi pada mamalia sebagian besar difokuskan pada homolog dari kandidat ini. . Selain itu, banyak dari kandidat ini hadir dalam mechanoreceptors kulit dan neuron somatosensori (Gambar. 2).
Saluran Ion Penginderaan Asam
ASIC termasuk dalam subkelompok proton-gated dari keluarga saluran Na + degenerin epithelial.74 Tiga anggota keluarga ASIC (ASIC1, ASIC2 dan ASIC3) diekspresikan dalam mechanoreceptors dan nociceptors. Peran saluran ASIC telah diteliti dalam studi perilaku menggunakan tikus dengan target penghapusan gen saluran ASIC. Penghapusan ASIC1 tidak mengubah fungsi mechanoreceptors kulit tetapi meningkatkan sensitivitas mekanik aferen yang menginervasi usus.75 Tikus KO ASIC2 menunjukkan penurunan sensitivitas LTMR kulit yang beradaptasi dengan cepat.76 Namun, penelitian selanjutnya melaporkan kurangnya efek melumpuhkan ASIC2 pada baik mechano-nociception visceral dan mechanosensation kulit.77 Gangguan ASIC3 menurunkan sensitivitas mechano aferen viseral dan mengurangi respons HTMR kulit terhadap rangsangan berbahaya.76
Saluran Resep Transien
Superfamili TRP terbagi menjadi enam subfamili pada mamalia.78 Hampir semua subfamili TRP memiliki anggota yang terkait dengan mekanosensasi dalam berbagai sistem sel.79 Pada neuron sensorik mamalia, saluran TRP paling dikenal untuk mendeteksi informasi termal dan mediasi inflamasi neurogenik, dan hanya dua saluran TRP, TRPV4 dan TRPA1, telah terlibat dalam respons sentuh. Mengganggu ekspresi TRPV4 pada tikus hanya memiliki efek sederhana pada ambang mekanosensori akut, tetapi sangat mengurangi kepekaan terhadap rangsangan mekanis berbahaya. 80,81 TRPV4 adalah penentu penting dalam membentuk respon neuron nociceptive terhadap stres osmotik dan hiperalgesia mekanik selama inflamasi.82,83 TRPA1 tampaknya memiliki peran dalam hiperalgesia mekanik. Tikus yang kekurangan TRPA1 menunjukkan rasa sakit hipersensitivitas. TRPA1 berkontribusi pada transduksi rangsangan mekanis, dingin dan kimia pada neuron sensoris nociceptor tetapi tampaknya tidak penting untuk transduksi sel rambut.84,85
Tidak ada bukti yang jelas yang menunjukkan bahwa saluran TRP dan saluran ASIC yang diekspresikan pada mamalia secara mekanis dikunci. Tak satu pun dari saluran ini menyatakan secara heterolog merekapitulasi tanda tangan listrik dari arus mekanositif yang diamati di lingkungan asli mereka. Ini tidak mengesampingkan kemungkinan bahwa ASIC dan saluran TRPs adalah mekanotransduser, mengingat ketidakpastian apakah saluran mekanotransduksi dapat berfungsi di luar konteks seluler (lihat bagian pada SLP3).
Protein Piezo
Piezo protiens baru-baru ini diidentifikasi sebagai kandidat yang menjanjikan untuk protein mechanosensing oleh Coste dan kolaborator. Vertebrata 86,87 memiliki dua anggota Piezo, Piezo 1 dan Piezo 2, sebelumnya dikenal sebagai FAM38A dan FAM38B, masing-masing, yang dilestarikan dengan baik di seluruh eukariota seluler. Piezo 2 melimpah di DRG, sedangkan Piezo 1 hampir tidak terdeteksi. Piezo-induced mechanosensitive current dicegah dihambat oleh gadolinium, ruthenium red dan GsMTx4 (racun dari tarantula Grammostola spatulata) .88 Ekspresi Piezo 1 atau Piezo 2 dalam sistem heterolog menghasilkan arus mechanosensitive, kinetika inaktivasi Piezo 2 saat ini menjadi lebih cepat. dari Piezo 1. Mirip dengan arus mekanositif endogen, arus yang bergantung pada Piezo memiliki potensi pembalikan di sekitar 0 mV dan kation tidak selektif, dengan Na +, K +, Ca2 + dan Mg2 + semuanya menembus saluran yang mendasarinya. Demikian pula, arus yang bergantung pada piezo diatur oleh potensial membran, dengan perlambatan kinetika arus yang jelas pada potensi yang terdepolarisasi.86
Protein piezo tidak diragukan lagi adalah protein yang meresepkan protein dan berbagi banyak sifat dari arus mekanositif yang cepat beradaptasi di neuron sensorik. Pengobatan neuron DRG berbudaya dengan Piezo 2 mengganggu RNA mengurangi proporsi neuron dengan cepat beradaptasi saat ini dan penurunan persentase neuron mechanosensitive. Domain transmembran 86 terletak di seluruh protein piezo tetapi tidak ada motif yang mengandung pori yang jelas atau tanda saluran ion telah diidentifikasi. Namun, protein Piezo 1 tikus dimurnikan dan disusun kembali menjadi bilayers lipid asimetris dan liposom membentuk saluran ion yang sensitif terhadap ruthenium merah. 87 Langkah penting dalam memvalidasi mekanotransduksi melalui saluran Piezo adalah dengan menggunakan pendekatan in vivo untuk menentukan kepentingan fungsional dalam sinyal sentuh. Informasi diberikan di Drosophila di mana penghapusan anggota Piezo tunggal mengurangi respon mekanis terhadap rangsangan berbahaya, tanpa mempengaruhi sentuhan normal. 89 Meskipun struktur mereka masih harus ditentukan, keluarga baru protein mechanosensitive ini adalah subjek yang menjanjikan untuk penelitian masa depan, di luar perbatasan. sensasi sentuhan. Sebagai contoh, penelitian terbaru pada pasien dengan anemia (xerositosis herediter) menunjukkan peran Piezo 1 dalam mempertahankan homeostasis volume eritrosit. 90
Transmembrane Channel-Like (TMC)
Sebuah studi baru-baru ini menunjukkan bahwa dua protein, TMC1 dan TMC2, diperlukan untuk mekanotransduksi sel rambut. 91 Ketulian herediter karena mutasi gen TMC1 dilaporkan pada manusia dan tikus. 92,93 Kehadiran saluran ini belum ditunjukkan dalam sistem somatosensori, tetapi tampaknya menjadi petunjuk yang bagus untuk diselidiki.
Stomatin-Like Protein 3 (SLP3)
Selain saluran transduksi, beberapa protein aksesori yang terhubung ke saluran telah terbukti memainkan peran dalam kepekaan sentuhan. SLP3 dinyatakan dalam neuron DRG mamalia. Studi menggunakan tikus mutan yang kurang SLP3 telah menunjukkan perubahan dalam mekanosensasi dan arus mekanosatif. Fungsi akurat 94,95 SLP3 masih belum diketahui. Ini mungkin menjadi penghubung antara saluran mechanosensitive dan mikrotubulus yang mendasari, seperti yang diusulkan untuk C. elegans homolog MEC2.96 GR. Lab Lewin telah menyarankan bahwa penambat disintesis oleh neuron sensorik DRG dan menghubungkan saluran ion mechanosensitive ke matriks ekstraseluler. 97 Mengganggu tautan menghapuskan arus RA-mechanosensitive yang menunjukkan bahwa beberapa saluran ion hanya mekanifitif ketika ditambatkan. Arus RA-mechanosensitive juga dihambat oleh laminin-332, protein matriks yang diproduksi oleh keratinocytes, memperkuat hipotesis modulasi arus mekanosensitif oleh protein ekstraseluler.98
K + Saluran Subfamili
Sejalan dengan kationic depolarizing mechanosensitive currents, kehadiran repolarizing mechanosensitive K + current sedang diselidiki. Saluran K + dalam sel yang sangat sensitif dapat melangkah dalam keseimbangan saat ini dan berkontribusi untuk menentukan ambang mekanik dan waktu adaptasi mekanoreptor.
Anggota KCNK termasuk dalam domain K + channel dua-pori (K2P) family.99,100 K2P menampilkan serangkaian regulasi yang luar biasa oleh agen seluler, fisik dan farmakologis, termasuk perubahan pH, panas, peregangan dan deformasi membran. K2P ini aktif pada potensial membran istirahat. Beberapa subunit KCNK dinyatakan dalam neuron somatosensori. 101 KCNK2 (TREK-1), KCNK4 (TRAAK) dan saluran TREK-2 adalah di antara beberapa saluran di mana gating mekanis langsung dengan peregangan membran telah ditunjukkan.102,103
Tikus dengan gen KCNK2 yang terganggu menunjukkan sensitivitas yang ditingkatkan terhadap panas dan rangsangan mekanis ringan tetapi ambang penarikan normal terhadap tekanan mekanis berbahaya yang diterapkan pada kaki belakang menggunakan uji Randall Selitto.104 Tikus yang kekurangan KCNK2 juga menunjukkan peningkatan hiperalgesia termal dan mekanis pada inflamasi kondisi. Tikus knockout KCNK4 hipersensitif terhadap stimulasi mekanis ringan, dan hipersensitivitas ini meningkat dengan inaktivasi tambahan KCNK2.105 Peningkatan mekanosensitivitas dari tikus knockout ini dapat berarti bahwa regangan biasanya mengaktifkan arus mekanositif depolarisasi dan repolarisasi secara terkoordinasi, mirip dengan ketidakseimbangan depolarisasi dan repolarisasi arus gerbang tegangan.
KCNK18 (TRESK) adalah kontributor utama konduktansi K + latar belakang yang mengatur potensi membran istirahat dari neuron somatosensori.106 Meskipun tidak diketahui apakah KCNK18 secara langsung sensitif terhadap stimulasi mekanis, ia mungkin berperan dalam memediasi respons terhadap sentuhan ringan, serta rangsangan mekanis yang menyakitkan. KCNK18 dan pada tingkat yang lebih rendah KCNK3, diusulkan untuk menjadi target molekuler dari hidroksi -? - sanshool, senyawa yang ditemukan dalam biji merica Schezuan yang mengaktifkan reseptor sentuhan dan menyebabkan sensasi kesemutan pada manusia.107,108
Saluran K + yang bergantung pada tegangan KCNQ4 (Kv7.4) sangat penting untuk mengatur kecepatan dan preferensi frekuensi dari subpopulasi mekanisme mekanis yang cepat beradaptasi pada tikus dan manusia. Mutasi KCNQ4 awalnya dikaitkan dengan bentuk tuli herediter. Menariknya, penelitian terbaru melokalisasi KCNQ4 di ujung saraf tepi kulit dengan cepat mengadaptasi folikel rambut dan Meissner corpuscle. Dengan demikian, hilangnya fungsi KCNQ4 mengarah pada peningkatan sensitivitas mekanoreptor yang selektif terhadap getaran frekuensi rendah. Khususnya, orang-orang dengan gangguan pendengaran akhir-onset karena mutasi dominan dari gen KCNQ4 menunjukkan peningkatan kinerja dalam mendeteksi amplitudo kecil, getaran berfrekuensi rendah.109

Wawasan Dr. Alex Jimenez
Sentuhan dianggap sebagai salah satu indra paling kompleks dalam tubuh manusia, terutama karena tidak ada organ khusus yang bertanggung jawab atasnya. Sebaliknya, indera peraba terjadi melalui reseptor sensorik, yang dikenal sebagai mechanoreceptors, yang ditemukan di kulit dan merespon tekanan mekanis atau distorsi. Ada empat jenis utama dari mechanoreceptors pada kulit mamalia yang gundul, atau tidak berbulu: selsel lamellar, corpuscles taktil, ujung saraf Merkel dan sel-sel bulat. Fungsi mechanoreceptors untuk memungkinkan deteksi sentuhan, untuk memantau posisi otot, tulang dan sendi, yang dikenal sebagai proprioception, dan bahkan untuk mendeteksi suara dan gerakan tubuh. Memahami mekanisme struktur dan fungsi mekanoreptor ini adalah elemen mendasar dalam pemanfaatan perawatan dan terapi untuk manajemen nyeri.
Kesimpulan
Sentuhan adalah rasa yang kompleks karena mewakili kualitas taktil yang berbeda, yaitu, getaran, bentuk, tekstur, kesenangan dan rasa sakit, dengan pertunjukan diskriminatif yang berbeda. Hingga kini, korespondensi antara organ-sentuhan dan rasa psikofisik adalah penanda molekuler yang bersifat korelatif dan kelas-spesifik baru saja muncul. Pengembangan tes hewan pengerat yang sesuai dengan keragaman perilaku sentuhan sekarang diperlukan untuk memfasilitasi identifikasi genom masa depan. Penggunaan tikus yang tidak memiliki himpunan bagian tertentu dari jenis aferen sensorik akan sangat memudahkan identifikasi mekanoreptor dan serabut aferen sensoris yang terkait dengan modalitas sentuhan tertentu. Menariknya, sebuah makalah baru-baru ini membuka pertanyaan penting dari dasar genetik sifat mekanosensori pada manusia dan menunjukkan bahwa mutasi gen tunggal dapat mempengaruhi sensitivitas sentuhan negatif. 110 Ini menggarisbawahi bahwa patofisiologi defisit sentuhan manusia dalam bagian besar yang tidak diketahui dan pasti akan kemajuan dengan mengidentifikasi secara tepat bagian-bagian neuron sensori yang terhubung dengan modalitas sentuhan atau defisit sentuhan.
Sebagai imbalannya, kemajuan telah dibuat untuk menentukan sifat biofisik dari arus mechano-gated.64 Pengembangan teknik baru dalam beberapa tahun terakhir, memungkinkan pemantauan perubahan tegangan membran, saat merekam arus mechano-gated, telah terbukti metode eksperimental yang berharga untuk menggambarkan arus mechanosensitive dengan adaptasi cepat, menengah dan lambat (ditinjau dalam Delmas dan kolaborator) .66,111 Masa depan akan menentukan peran properti saat ini dalam mekanisme adaptasi mekanoreceptor yang beraneka ragam dan kontribusi arus K + yang mekanosensitif terhadap rangsangan LTMR dan HTMR.
Sifat molekuler arus mechano-gated pada mamalia juga merupakan topik penelitian yang menjanjikan di masa depan. Penelitian masa depan akan berkembang dalam dua perspektif, pertama untuk menentukan peran molekul aksesori yang menambatkan saluran ke sitoskeleton dan akan diperlukan untuk memberikan atau mengatur mekanosensitivitas saluran ion dari sejenis TRP dan ASIC / EnaC keluarga. Kedua, untuk menyelidiki area besar dan menjanjikan dari kontribusi saluran Piezo dengan menjawab pertanyaan kunci, relatif terhadap mekanisme permeasi dan gating, subset neuron sensorik dan modalitas sentuhan yang melibatkan Piezo dan peran Piezo dalam sel non neuronal yang terkait dengan mechanosensation.
Indra peraba, dibandingkan dengan indra penglihatan, pengecap, suara, dan penciuman, yang menggunakan organ tertentu untuk memproses sensasi ini, dapat terjadi di seluruh tubuh melalui reseptor kecil yang dikenal sebagai mechanoreceptors. Berbagai jenis mechanoreceptors dapat ditemukan di berbagai lapisan kulit, di mana mereka dapat mendeteksi berbagai macam rangsangan mekanis. Artikel di atas menjelaskan sorotan khusus yang menunjukkan kemajuan mekanisme struktural dan fungsional dari mechanoreceptors yang terkait dengan indra peraba. Informasi yang dirujuk dari Pusat Nasional untuk Informasi Bioteknologi (NCBI). Cakupan informasi kami terbatas pada chiropraktik serta cedera dan kondisi tulang belakang. Untuk membahas pokok bahasan ini, jangan ragu untuk bertanya kepada Dr. Jimenez atau hubungi kami di 915-850-0900 .
Diundangkan oleh Dr. Alex Jimenez

Topik Tambahan: Back Pain
Nyeri punggung adalah salah satu penyebab utama kecacatan dan hari-hari yang terlewatkan di dunia kerja. Nyatanya, nyeri punggung telah dianggap sebagai alasan paling umum kedua untuk kunjungan ke dokter, hanya kalah jumlah oleh infeksi saluran pernapasan atas. Sekitar 80 persen populasi akan mengalami beberapa jenis nyeri punggung setidaknya sekali sepanjang hidup mereka. Tulang belakang adalah struktur kompleks yang terdiri dari tulang, sendi, ligamen dan otot, di antara jaringan lunak lainnya. Karena ini, cedera dan / atau kondisi yang diperburuk, seperti cakram hernia, akhirnya dapat menyebabkan gejala nyeri punggung. Cedera olahraga atau cedera kecelakaan mobil sering menjadi penyebab paling sering dari nyeri punggung, namun terkadang gerakan yang paling sederhana dapat memiliki hasil yang menyakitkan. Untungnya, pilihan pengobatan alternatif, seperti perawatan chiropractic, dapat membantu meringankan nyeri punggung melalui penggunaan penyesuaian tulang belakang dan manipulasi manual, yang pada akhirnya meningkatkan pereda nyeri.

TOPIK EXTRA PENTING: Manajemen Nyeri Punggung Rendah
TOPIK LAINNYA: EKSTRA EKSTRA: Perawatan & Perawatan Kronis
Kosong
Referensi
Tutup Akordeon
Informasi di sini tentang "Mekanisme Struktural dan Fungsional dari Mechanoreceptors" tidak dimaksudkan untuk menggantikan hubungan pribadi dengan profesional perawatan kesehatan yang berkualifikasi atau dokter berlisensi dan bukan merupakan saran medis. Kami mendorong Anda untuk membuat keputusan perawatan kesehatan berdasarkan penelitian dan kemitraan Anda dengan profesional perawatan kesehatan yang berkualifikasi.
Informasi Blog & Ruang Lingkup Diskusi
Lingkup informasi kami terbatas pada Chiropractic, musculoskeletal, obat-obatan fisik, kesehatan, kontribusi etiologis gangguan viscerosoma dalam presentasi klinis, dinamika klinis refleks somatovisceral terkait, kompleks subluksasi, masalah kesehatan sensitif, dan/atau artikel, topik, dan diskusi kedokteran fungsional.
Kami menyediakan dan menyajikan kerjasama klinis dengan para ahli dari berbagai disiplin ilmu. Setiap spesialis diatur oleh ruang lingkup praktik profesional mereka dan yurisdiksi lisensi mereka. Kami menggunakan protokol kesehatan & kebugaran fungsional untuk merawat dan mendukung perawatan cedera atau gangguan pada sistem muskuloskeletal.
Video, postingan, topik, subjek, dan wawasan kami mencakup masalah, masalah, dan topik klinis yang terkait dengan dan secara langsung atau tidak langsung mendukung ruang lingkup praktik klinis kami.*
Kantor kami telah berusaha secara wajar untuk memberikan kutipan yang mendukung dan telah mengidentifikasi studi penelitian yang relevan atau studi yang mendukung postingan kami. Kami menyediakan salinan studi penelitian pendukung yang tersedia untuk dewan pengawas dan publik atas permintaan.
Kami memahami bahwa kami mencakup hal-hal yang memerlukan penjelasan tambahan tentang bagaimana hal itu dapat membantu dalam rencana perawatan atau protokol perawatan tertentu; oleh karena itu, untuk membahas lebih lanjut materi pelajaran di atas, jangan ragu untuk bertanya Dr Alex Jimenez, DC, atau hubungi kami di 915-850-0900.
Kami di sini untuk membantu Anda dan keluarga Anda.
Berkah
Dr. Alex Jimenez IKLAN, MSACP, RN*, CCST, IFMCP*, CIFM*, ATN*
email: pelatih@elpasofungsionalmedicine.com
Lisensi sebagai Doctor of Chiropractic (DC) di Texas & New Mexico*
Lisensi Texas DC # TX5807, Lisensi New Mexico DC # NM-DC2182
Berlisensi sebagai Perawat Terdaftar (RN*) in Florida
Lisensi Florida Lisensi RN # RN9617241 (Kontrol No. 3558029)
Status Kompak: Lisensi Multi-Negara: Berwenang untuk Praktek di Status 40*
Alex Jimenez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
Kartu Bisnis Digital Saya





